Reproduction in Plant, Biology, Class 12 PDF Download
Introduction and Flower parts - Sexual Reproduction in Flowering Plants, Biology, Class 12
LIFE CYCLE OF ANGIOSPERMS (CAPSELLA)
CLASSIFICATION
Class - Dicotyledonae
Subclass - Polypetalae
Series - Thalamiflorae
Order - Parietales
Family - Cruciferae or Brassi caceae
Genus- Capsella
Species - bursa pastoris [Common name "Shepherd's purse" ]
SPECIAL POINTS TO BE REMEMBER
-
Angiosperm orginated in Mesozoic era .
-
Angiosperm originated either in the begining of Cretacious period or in ending of Jurassic period of Mesozoicera . It means they are originated between Cretacious and Jurassic period on the earth.
-
Angiosperm dominated over the earth in Coenozoicera . Sothisera is known as "Golden Period of Angiosperms" .
-
First of all N.Grew realized the fact, Stam ens are male sex organ of flower (Anatomy of plants)
-
Sexuality in plant first of all reported by Jacob Camerarius .
-
He reported Anthers are the male sex organ and Ovary with style and stigma are female sex organ , and for the formation of seed, interaction is essential in between both the sex organs.
-
Significance of pollination and role of insects in pollination was recognized by Josheph Kolreuter .
-
C.F.Wolf - Father of plant Embryology.
-
Prof. P. Maheshwari – Father of Indian plant Embryology. He wrote a book – 'An Introduction to
Embryology of Angiosperms'.
GENERAL INTRODUCTION
Most of the important Angiospermic characters are found in Capsella so that for the study of Angiosperms, it is considerd as a "Typical Angiosperm".
-
It is an annual plant and grows as weed during the winter season in the field.
-
The main plant of the Capsella is a sporophyte. Which is diploid and it is differentiated into root, stem and leaves .
-
Capsella is a heterosporous plant it means there are two different types of spores are formed in the life cycle which is classified into two categories in which male spores are called Microspores and female spores are called Megaspores .
-
The proce ss of reproduction takes place in this plant through a special structure, called flower .
-
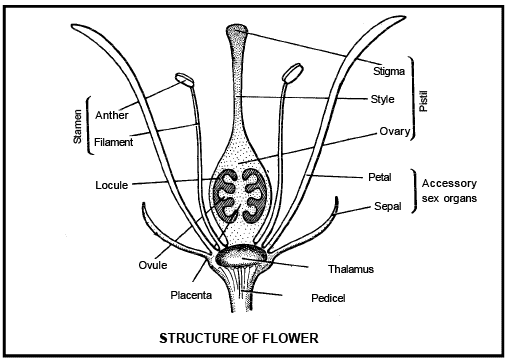
Calyx , Corolla , Androecium and Gynoecium are present in the typical or complete flower.
-
The calyx and corolla are termed accessary whorls of the flower, because these structures do not participate in the process of reproduction, only helps
-
The androecium and gynoecium are known as essential whorls , because they are directly related with the reproduction.
FLOWER IS A MODIFIED SHOOT
According by Goe the , Flower is modified shoot which shows favourable adaptation for reproduction through a special method. Flower has a small or long stalk like – structure called pedicel. Free end of the pedicel is flattaned or dome shaped is called thalamus . The thalamus is a type of odified stem , on which nodes and internodes are present.
-
Nodes are present very close to each other and internodes are small highly reduced in the thalamus.
-
The whorls present in the flower are the modifications of leaves and arranged in four circles on the thalamus.
-
The four nodes are present on the thalamus, in which first modified leaves are attached on the first lowernode are called calyx .
-
The corolla born on the second node . Androeci um are present on the third node and gynoecium on the fourth node in uppermost position.
-
In some of the plants the length of internode increases which is present in between Calyx and Corol la is called Anthophore e.g. Silene plant, Dianthus .
-
The length of internode between the corolla and androecium increases. It is called androphore e.g. Passiflora .
-
If the internode between androecium and gynoec ium increases then it is called gynophore . e.g. Capparis .
-
Both androphore and gynophore are present in the same flower is called gynandrophore or androgynophore . e.g. Gynandropsis pentaphylla and Cleome gynandra.
-
Calyx is also modifie d vegetative leaves . Such as in Mussaenda flower, one sepals of calyx modified into leaf like bright and attractive yellow colour structure called "Advertising flag". It helps in pollination
-
In some flowers; thalamus grows inside the ovary is called carpophore e.g. Coriandrum and Foeniculum .
-
In Nym phae a Petals like stam ens are pres ent.
-
On the basis of above examples , we can prove " Flower is modified shoot".
MONOCARPIC PLANTS :
-
The plants in which flowering and fruiting takes place only once in the whole life cycle are called monoc arpic e.g. Annual & Biennial plants.
POLYCARPIC PLANTS :
-
The plants in which flowering and fruiting takes place m an y times in the entire life cycle are known as polycarpic e.g. Perennial plant.
EXCEPTION :
Bamboo , Palms, Banana, Centuary plant (Agave americana) are perennial plants but they are the example of monocarpic plants.
REPRODUCTION – IN FLOWERING PLANTS - [EMBRYOLOGY]
"Reproduction is one of the important processes by which every living organism make a copy of itself. It is the means of multiplication and perpetuation of species because the older individual of each species undergo senescence and die"
All the reproductive methods of plants are broadly categorised into tw o types -
1. Sexual Reproduction
2. Asexual Reproduction
SEXUAL REPRODUCTION
In Ang i osp erms ma le an d f em ale g ame tes a re f orm ed in m ale and fe male sex organs by the proc ess of meiosis . Both the gametes fuse together to form a diploid zyg ote which gives rise embryo . The proces s in which embryo is formed by meiosis and fertilization is called Amphimixis.
Male Reproductive Organ – Androecium
Male reproductive organ is called androecium and their unit is called stamen.
Stamen is also known as mic ros porophyll. There are 6 stamens in Caps ella .
A typical stamen is differentiates into three parts -a long, thin structure is called f il a me n t which joins the stamen to the thalamus. The free end of the filament, a swollen spore bearing structure is called a nther. Anther and filament are attached together with help of small region, called co nnec tiv e. Connective contains va scular t issues . The main parts of the stamen is the anther .
Each an ther genera lly bilobed structure i.e ., anthe r has t wo an ther lobes is c alled dit heco us .
Each lobe of a nther h as two chamb ers whic h are cal led po llen s ac s or pollen chambers .

-
Therefore, a typical anther has four pollen sacs is called tetrasporang iate .
-
Pollen grains are formed inside the pollen sac through the meiotic division in pollen mother cells.
-
At the ma turity of the pollen grains, steri le tiss ue degenerate which are present in between the pollen sacs. Both the pollen sac fused together. Because of this reason, only one chamber appears in each anther lobe at maturity. So two chambers are seen in the mature anther at the time of dehisc ence.
-
In C a pse l la , whic h i s me mber of the c ru c if e ra e or Brassic ace ae , an ther ar e di the cous and tetra s poran g ia te type.
-
But in Malvac eae, the anther of stamen has only one anther l obe . Thi s is called monothecou s and it co ntai ns on ly t w o pol l en sa cs ca lled B is por an g ia te .
-
M on ot hec ou s a n t her are also found in M ori nga , W olffia pl ants.
-
In Arc euth obium where there is only one micros pora ngiu m per anther. This condition is called M on os p or an g ia te .

[ A ] STRUCTURE OF ANTHER :
The development of anth er in orig in is Eusp orangia te type i .e. it is d eveloped f rom more th en one archesporial cells.
-
In the transverse sect ion of anther, it is seen almost spherical.
-
The following structures are present in the anther :-
( i ) Epidermis :- It is t he outermost layer of a nt her. It is si ngle cel led t h ick a nd cont i nuous layer but not arche sporial i n origi n. It forms t he outermost protect ive layer .
-
In Ar ceuthobium (Smallest Parasitic Angiosperm) fibrous thickening present in epidermis so it is c alled exothecium.
(ii) Endot hec ium :- This layer is pre sent below the epidermis. It is single celled thick layer. During the maturat ion of anther, various changes takes place in different walls of cells of endothecium. The outer wall of these cells remains thin walled, but inner walls and radial walls become thick due to thickening of a–cellulose fibers.
Callose bands are also present along the radial walls. At some places callose bands and fibrous thickening are a bsent. Th ese place s are called st omium . The dehi sc ence of anth er takes palc e only from these places. En do the ci um becomes hygrosc opic nature due to presence of fibrous thickening. So it helps in dehiscence of anther.
Note: In Hydrocharitaceae family fibrous thickening is absent in endo thecium.
(iii) Middle layer :- Middle layer consists of parenchymatous cells.This layer is one to three cel led t hick str ucture . Food is s tore d by pa re nc h yma tous cells in this layer. Middle layer is ephemeral in nature and absent in a mature anther.

Note :
(1) In Hol optel ia plant 3 to 4 cel l ed thick middle layer is present.
(2) In Na jadac eae & Lemnaceae families middle layer is absent.
(3) In Wolffia middle layer is absent.
(iv) Tapet um :- I t is t he i nner most l ayer wh ich act s as nutritive layer. Pollen sacs are surrounded by tapetum.
This is also single celled thick layer. The cells of the tapetum initially dipl oid b ut t h ey become polyploid and mu ltin ucl ea te due to endomitosis, fr ee n uc le ar division and polyteny. It means these cells contain many chromosomes.
Tapetum ab sorbs foo d from the middle layer a nd provide nutrition to the microspore mother cel ls or microspores . The cells of tapetu m secrete hormo nes and enzymes . The tap etum layer disap pears in the mature anther.
N OT E : In N ic od ia and Cost um pla nts, tapetum is multilayer ed.
TAPETUM IS OF TWO TYPES :-
( i ) AMOEBOIDTAPETUM/ IN VASIVE TAPETUM/PERIPL AS MO DIAL TAPETUM : -It is found i n pr i mit ive Ang iospe rm . Such t ype of tapetum ab sorb a l l foods fro m t he m i dd l e l ay er. S o middle layer immediately degenerates. In the beginning, all food materials stored by tapetum. Tapetal cells convert absorbed food into spec ial food g ranules called protoplast bodie s . The innermost layer of tapetum dissolve and release its protoplast into the cavity of the microsporangium. Now inside the pollen sacs protoplast bodies are know n as pe riplas m odium. Microspore mother cells are surrounded by periplasmodium and provides nourishment to the developing microspores. This type of tapetum provide nutrition to the microspores after degeneration .
Ex ample : Typh a, Alis ma and Trade sc a nt ia .
(II) GLAN D U LAR O R S ECRETORY TAPETUM : -
It is ad van ced typ e of tapetum. It does not degenerates quickly. It absorbs nutrients from the middle layer and secretes into the cavity of the microsporangia (Pollen sacs) and does not store it e.g. Usually it is found in most of Flowe ring plants (Capsella) .
Before degeneration of cells of tapetum, they form special granules called Proubisc h bodies in cyt oplasm.
Proubisch bodies transfer between cell wall and cell membrane of tapetal cells. Here they are surrounded by s p orop ol leni n . Now they a re called Ub is ch bodi es or orb icules . At l as t ta pet um d eg en er ate s an d ub is ch bodies released into pollen sacs.
Generally, s poropollenin partic ipa tes in the formation of outer covering (Exine) of Pollen grains.
Note - Tapetum helps i n trans fe r of food, sto rage of food, format ion of sporopollenin and poll enkitt materials.
Poll ensacs : Four Pollen sac s are pre sentin the a nt her. Poll en s acs are also known as micro sporangia .
Ins ide the pollen s acs, microspores are fo rmed by the me iotic di vis ion of microspore mother cells .
===============================================================
Microsporogenesis - Sexual Reproduction in Flowering Plants, Biology, Class 12
[B] MICROSPOROGENESIS :
- The anther appears as outgrowth like structure in the initial stage which shows spherical or oval shaped structure.
- At this stage, it is a mass of undifferentiated and homogenous meri stematic cells whic h is surrounded by a single cell thick outer layer. This layer is known as epidermis. First of all vascular tissue are formed in middle region. Simultaneously group of cells located just below the epidermis in vertical rows in the region of hypodermis at the four corners are become large has visible nucleus with dense cytoplasm. Due to this reason they are different from the rest of the cells. These cells are called arches porial cells .
- These cells divide periclinally to form primary parietal cells below the epidermis and primary sporogenous cells towards the centre. Both of the cells usually undergo further divisions to form complete structure of anther except epidermis.
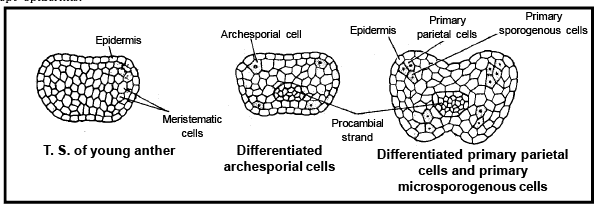
- Primary parietal cells undergo further periclinal and anticlinal division to forma serie s of 3-5 la yers making the walls of the anther.
- Out of them outer most layer of anther is formed just below the epidermis by primary parietal cells is called endothecium or fibrous layer . T he endothecium is fol lowed by 1-3 cel led th ick layer is termed middle layer . The i nn ermo st lay er of the anther which surrounds pollen sacs, is called tapetum . Later the tapetal cells play a significant role during the meiotic cell division in microsporogenous cells and in pollen development.
- The primary sporogenous cells divide twice or more than two by mitotic division to form sporogenous cells and later sporogenous differentiated into microspore mother cells during the formation of wall of pollensac.
- Each microspore mother cell div ide to form four haploid microspore or pollen grain by meiotic division or reduction division .
- During this period spherical bodies are formed inside the tapetal cells before their disintegration. These spherical bodies are known as Ubisch-body . Ubischbody is made up of a complex substance called sporopollenin . It is the polym er of carotenoids .
- After the formation of ubisch body, the tapetum layer degenerates. Ubisch bodies participate in the formation of exine of the microspores inside the pollen sacs. Now thick walled microspores are called pollen grains .
- At the init ial stage all four microspores are at tached toget her with the help of callose layer. T h is group of microspores is called tetrad. After some time, this callose layer dissolve by callase enzyme. Which is secreted by tapetum.
Not e :- Normally each micros pore mother cell can formtetrad by meiotic division. But in some plant like Zostera, some microspore mother cells become sterile and provide nutrition to rest of microspore mother cells.
Note Similarly, tapetum is not well developed in Gentianaceae family so some cells of sporogenous tissue become sterile and provide nutrition to remaining sporogenous cells.
Types of tetrads :- The arrangment of the microspores in tetrad condition as follows
(i) Tetrahedral tetrad :- Four haploid microspores arranged in tetrahedral form
Example :- Dicotyledons- (Capsella).
(ii) Isobilateral tetrad :- This condition is found in monocotyledons. Microspores are arranged at the lateral side of each other.
(iii) Decussate Tetrad :- In this two microspores lies at the right angle of other microspores
Example :- Magnolia
(iv) T-Shaped tetrad :- Two microspores lie longitudinally and two microspores lie at transversly in this type of tetrad. Example :- Aristolochia & Butomopsis.
(v) Linear tetrad - In this tetrad all four pollens arranged in linear order. e.g., Halophylla, Halophia.
- All the above type of tetrads are found in Aristolochia elegans.
- Most common type of tetrads is Tetrahedral.


=================================================
Pollen grains - Sexual Reproduction in Flowering Plants, Biology, Class 12
Facts about types of Pollen grains –
(1) In Ascl epiadace ae (Calotropis ) and Orchidaceae family, all the pollen grains joined together to fo rm " Polli nium ". Pollinium of Calotropis is also called "T rans lator apparatus" .
( 2 ) More than four pollen grains are found intetrad called " Polyspory " e.g. Cuscuta .
( 3 ) In some plants, four pollens of tetrad join together permanently are called "Compound pollens " e.g.
Drosera, Typha, Drimys and Elodea .
(4) Compound pollens of Mimosaceae (Mimosa) family contains 6-8 or 64 pollen grains stick together to form a small unit that is cal led " Massullae " e.g. Mimosa and Neottia plants.
(5) Pollengrai ns of some plants present in air cause al lergy are called "aeroallergens " e.g.
Chenopodium, Parthenium, Sorghum , and Am aranthus . ["Hay fever" is caused by pollens of Ambr sia .]
( 6 ) Pollen grains of many plants are rich in nutrients. It has becomes a fashion in recent years to use pollen tabletes as food supplements. In western countries, a large number of pollen products in the form of tablets and syrups are available in the market. Pollen consumption has been claimed to increase the performance of athletes and race horses.
( 7) In Cypera ceae family only one pollen grain is formed from pollen mother cell. e.g., Cyperus .
(8) Largest pollen - Mirabilis .
(9) Smallest pollen - Myosotis.
(1 0) Longest pollen – Zostera (Fili form pollen)
(11) Eight nucleated embryo s ac type of po llen is found in Hyacinthus .[T his type of pollen grain discovered by Nemec . So it is called Nemec phenomenon ]
( 1 2) Dumble shaped pollens are found in Vincaherbacea .

[ C ] STRUCTURE OF M ICROSPORE OR POLLEN GRAIN :

Pollen grain is the first cell of a male gametophyte.
- Pollen grain is termed as immature male g ametophyte .
Usually, they are in round shape. Pollen grain surrounded by two d istinct la yers . The outer layer (wall ) is thick, rigid and orna mented , called exine . Thi s l aye r i s formed by c utin and s porop ollenin . Sporopollenin is highly re istent material . It is nonbiodegrada ble.
- Due to the presence of sporopollenin, fos sils o f pollen g rai n are always found in good condition . The presence of fossils of pollengrains can forecast the presence of natural reso urc e s like p et rol e um , coa ls e tc. in the eart h.
- The in ternal l aye r is thin, soft and el ast ic in nature. It is called intine. It is made up of pectin and Cellu lose or pecto- cellulose.
- Usually, at few place s on outer surface exine is absent or pre sent in the form of thin layer. The se thin place s are called germ pore. The intine comes out through the any one germpore during the germination of pollen grain in the form of pollen tube .
- The number of germpore , structure and orname ntat ion of ex ine is a sig nifi cant feature of taxonomy .
- A detail study of pollen grains is called Palynology.
- Three colpus type (slit type) of germpore are present in pollen grain of most of the dicots (Capsella) . This type of p ollen grains are called tricolpate . Only one germ pore is present in monocots and pollen g rain is called monocolpate.
- The plants in which pollination takes place by insects , their pollen grains having oily layer around the pollen grain. It is called pollen -kitt. It is composed of lipids and carotenoids.
Function of pollen kitt :-
(i) This oily layer protects the pollen grain from the harmful ultraviolet rays.
(ii) Its stic ky surface helps to attach with the insec ts.
(iii) Its yellow c olour attracts the insects. Pollen kit is pres ent on the pollens of Caps ella.


[D] DEHISCENCE OF ANTHER :
During the maturation of anther, various changes are takes place in walls of anther.
- In the beginning, middle layer degenerates due to absorpti
- When the micropores are formed inside the pollen sacs, at the same time ubisch bodies are formed in cells of tapetum, then after it degenerates. Ubisch bodies participate in the formation of exine of pollens.
- In this way, in a mature anther only two layers epidermis and endothecium are present in the form of outer covering.
- The sterile tissues are present between both the pollen sacs of each anther lobe degenerate. So both pollen sacs of the each anther lobe fuse together to form single pollen sac
- Therefore, in the T.S. of mature anther only two pollen sacs are present.
- Dehiscence of anther takes place during the dry season. Due to the hygroscopic nature of endothecium, loss of water takes place from the cell of endothecium.
- Walls of endothecial cells try to contract due to the loss of water but inner and radial walls do not contract due to presence of fibrous thickening whereas outer thin walled cells of endothecium contract and become concave or incurved.
- Incurving of outer walls exert pulling force or tension over the entire surface of anther. Due to tension, thin walled stomial cells breaks off and dehiscence of anther takes place and pollen grains are present in pollen sacs released into the atmosphere.
- Dehiscence of anther in Angiosperms either longitudinal or Apical pore, or Transverse or Valvular type. Dehiscence of anther of Capsella is longitudinal.


[ E ] MICRO- GAMETO GENESIS OR DEVELOPMENT OF MALE GAMETOPHYTE :
In flowering plants , pollen grain or mi cros pore considered as first cell of male gametophyte . Germination or development of pollen grain takes place before dehiscence of anther. So it is called as precocious development . Development of pollen also takes place at mother place [inside pollensac of anther] is called In-situ development.
( i ) Pre Pollinat ion devel op ment – In the beginning of the the process, only nucleus of pollen grain divided by unequal mitotic division , resulting two unequal s ize of nucleus are formed. Small nucleus present near the wall is called generative nucleus a nd large nucleus pre ent inside the cytoplasm is cal led Tube or Vegetat ive nucleus .
Both the nucleus surrounded by cytoplasm and it becomes dense, then followed by unequal cytokinesis , resulting two une qual size of the cells are formed. larger cell in which large nucleus is present known as Vegetative cell and smaller cell in which small nucleus is present , called generative cell .
Now pollen grains come in bicelled a nd binucle ated stage . In Angiosperms pollination of pollen grains take place in icelled and binucleated stage i n 60% A ngi ospe rms and in 40% An gio sperms pollination occurs at 3-celled stage. In this generative cell divides and form two male gametes. The development gametophyte take place inside the sporangia is also known as endosporic development .
This stage of pollen grain is called immature or partially developed male gam etophyte .
Generative cell detached from the wall and changed into vermiform or spindle shaped struc ture and enter inside the vegetative cell.
( i i ) Post Pollination development – Further develo ment of pollen grain [Immature male gemetophyte] takes place on the sti gma of Carpel after pollination. Pollens absorb moisture and sugar content from the stigma. Due to this volume of internal contents of cytoplasm increased. It exerts pressure on the both outer layers. Because of this pressure intine comes out through any one germp ore in the form of tube like st ructure called pollen tube .
First of all vegetative nucleus ent er into the p ollen tube and as sumes terminal [tips] position. Th is spindle shaped generative now enter into the pollen tube. Inside the pollen tube, generative cell divides mi tot ically and to form a two non m oti le ma le gametes . Now male gametophyte comes in three celled structure in which one vegetative cell and two male gametes are presen t.
This three celled stage represents the mature male gametop hyte of Angiosperm [Capsellaalso]. Mal e gam etophyte i s hi ghly reduc ed and c ompletely de pen ds on sporophy te.
First ofall pollen tube disc ove red by G.B. Amici in Portulaca plant .
Longest pollen tube is found in Zeamays .

==================================================================
FEMALE REPRODUCTIVE ORGAN-GYNOECIUM - Sexual Reproduction in Flowering Plants, Biology, Class 12
FEMALE REPRODUCTIVE ORGAN-GYNOECIUM
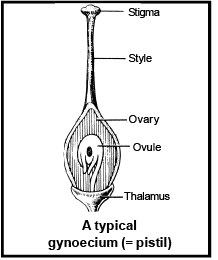
Gynoecium is the female reproductive organ. The free unit of gynoec ium is called pistil or carpel .
Carpel is also known as meg as porophyll .
The carpel is differentiate into three distinct region –
[i] Sti gma [ii] Style [iii] Ovary
The free end of the carpel which receives pollen grains is called stigma.
A long, narrow tubular structure is present in between the stigma and ovary called style. The basal swollen part of the carpel is called ovary.
The ovules is also known a s megas porongia which are borne on a cushion-like tis sue ca lled placenta in the ovary. One or more than one ovules are present inside the ovary.
The gynoecium of the Capsella is bicarpella y, syncar ous, unilocular and superior. It becomes bilocular due to the formation of false septum or replum at maturity.
[ A ] STRUCTURE OF OVULE OR MEGASPORANGIUM

A ridge or stalk like out growth is formed from the placenta of the ovary on which body of ovules are present.
Each ovule attached to the placenta by means of a thin stalk called funicle or funiculus/ Funiculum The point of attachment of the funicle with the ovule is called ilum .
The main region of the ovule is composed by mass of parenchymatous cells called nucellus . Nucellus is the main part of ovule . The nucellus is covered by one or two coats c alled integuments .
– In most of the ovule, funicle is attached to the main body of ovule for some distance (at lateral side) to form a ridge like struc ture k nown a s Raphe .
– Vascular tissue s are pre sent inside the funiculus which supply food material from the placenta to the body of ovule.
– A plac e from where funicle and integumen ts arise is called Chalaza .
Integument is absent just opposite to the chalaza, so that a narrow passage (pore) is formed which is called micropyle .
– icropyle in bitegmicovule has two pa rts - outer region w hich is surrounded by outer integument is Called exostome .
– The inner part of micropy le which is surrou nded by inner integument called end ostome .
– In most of the Angiosperm entire part of the nucellus is utilized by developing embryo sac but in some of the Angiosperm some part of the nuclellus remain inside the ovules that part of the nucellus present inside the seed in the form of a thin layer known as perisperm. Perisperm is commonly found in Piperaceae (Pipernigrum ) and Zingiberaceae Families (Turmeric, Ginger)
-
Some filaments are attached wi th funicl e [some times pl acenta] a re known as " Obturators " .
-
The function of obturators is to guide the passage of pollen tube towards the micropyle inside the ovary.
SPECIAL INTEGUMENTS –
(i) ARIL – It is the type of third integuments which develops from funicle at the base of the ovule e.g. Myristica , Asphodelus and Litchi.
(ii) ARILLODE – It develops from the tips of the outer integument and grows downwards and surrounds the entir e ovu le e. g. Pithe colobium (Inga dulce )
(iii) SARCOTESTA - When outer integument becomes fleshy then it is called sarcotesta e.g. Magnoliaceae (Magnolia).
(iv) OPERCULUM - It is a stopper or break like structure which is formed on the micropyle. It is formed due to the elongation of inner integuments. e.g. Lemnaceae family (Lemna).
(v) CARUNCLE OR STROPHIOLE - It is formed due to the proliferation (out growth) of outer integuments over the micropyle. e.g. Ricinus communis (Castor). It is formed by sugary contents so helps in absorption of water during germination of seeds and dispersal of seeds by ants called myrmecochory.
(vi) COMA - In some of the plants unicellular filaments like structures are present on the seed which is formed by cells of outer surface of outer integument. Such seeds are known as "Comose seeds". e.g.
Calotropis and Gossypium.
TYPES OF OVULE ON THE BASIS OF INTEGUMENTS :
A single integumented ovule is called unitegmic ovule - example - members of Gamopetalae and Gymnosperm.
Two integumented ovule is called bitegmic ovule. Example - In most of Angiosperm [Polypetalae-Capsella and Monocots].
-
The ovule in which integuments are absent is called Ategmic ovule e.g. Olax, Liriosma, Loranthus & Santalum.
TYPES OF OVULES ON THE BASIS OF NUCELLUS -
(i) Tenuinucellate - The nucellus is either less developed or present in the form of single layer.
Example :- Gamopetalae group.
(ii) Crassinucellate :- The nucellus is massive type i.e., it is made up of many layers.
Example :- Polypetalae group and Monocots
-
The nucellus degenerates in plants of Compositae family and integuments becomes active to form a nucellus like tissue. This is called endothelium or integumentary tapetum. It is polyploid structure.
-
The nucellus dissolves in the members of Podostemaceae family to form a nutritional cavity. This is termed pseudoembryo-sac
[ B ] TYPES OF OVULES :
There are six different types of ovules are found in Angiosperms on the basis of relationship of the micropyle, chalaza, and hilum with body of the ovule and orientation on the funiculus:
[ i ] AT ROPOUS OR ORTHOTROPOUS :-
The body of ovule is upright in position. The micropyle, chalaza and hilum lie in one straight line, so that this ovule is called straight or upright ovule. Example :- Betel, Piper, Polygonum and in Gymnosperms. It is the most primitive and most simple type of ovule of Angiosperms. Raphe is absent.

[ii] AN ATRO PO US OV ULE :-In this type, the body of the ovule completely turned at 1800 ang le, due to unilateral growth of funiculus, so it is also called inverted ovule. The chalaza and micropyle lie in straight line. The hilum and micropyle lie side by side very close to each ot her. Th is t ype of ovule is found i n 80% f a mi l ie s of Angiosperms but not in Caps e lla . In this ovule micropyle facing downward condition. This is the mos t comm on ty pe of ov ule so tha t it is co nsidered a s a "typic al ovule" of Ang iospe rms . eg. Members of Mal vace ae, Cuc u rb itac ea e, S olan ac ea e, C ompo si tae fa mi ly. I t i s als o cal le d res up ina te ov ul e.
[iii] HEMITRO POUSORHEMI - AN ATROPOUSOVULE :-In this ovule, the body of the ovule bent on funcile at 90 0 angle , i.e., body of ovule present at right a ng le to the funiculus. This is intermediate type between ortho and anatropous ovules. This ovule is also called ho ri zon ta l ovule because body of ovule present in horizontal position on the funiculus.
Micropyle and chalaza are present in the same line but micropyle is situated away from hilum. Example :-Ra nunculus , Primula, Golphimi a.
[iv] CAMPYLOTRO POUSOVULE :-In this ovule, the body of ovule curved (Curvature is not effective) in this way so micropyle and chalaza do no t prese nt in s tra ight li ne. The n uce llus i s p res ent in c u rve d pos iti on bu t the emb ryo s a c rema in straight. Micropyle comes c lose to the hilum . Eg.:- Leg u minosa e, Capparidac eae, C rucife rae fam ily [Ca pse l la]
[ v ] AMPH ITROPOUSOVULE :-In this type of ovule, curvature is more pronounced or effective in the nucellus and due to this effect of nuce llus, embryo sac becomes hors e shoe shape d. Micropyle comes close to the hilum. It is also called as transverse ovule. e.g. Mirabil is , Lemna , Poppy, Alisma, Buto maceae family.
[vi] CIRCIN OTROPOUSOVULE :-In this type of ovule, body of ovule inverted and again turned into straight position due to the growth of funiculus so that body of ovule present on funicle at 3600. The entire body of ovule is surrounded by funiculus.
It is also known as coiled ovule. Micropyle is situated away from hilum.e.g. Cactaceae family - O puntia .
===============================================================
Megasporogenesis and Female gamete formation - Sexual Reproduction in Flowering Plants, Biology, Class 12
[C] MEGASPOROGENESIS :
During the development of ovule, in the beginning of this process, nu ce ll us develops form the pl ac en ta in the form of a small rounded out growth like structure. At this stage, all the cells of nucellus are undifferentiated, homogenous and meristematic. This mass of cells surrounded by single celled thick layer of epidermis.

Any on e hy pode rmal cell of nucellus is differentiated and increase in size. It becomes different from rest of the cells due to presence of distinct nucleus. It is called arche sporial cell. Archesporium divides periclinally to form an oute r prima ry parie tal c ell and in ne r P rim ary S p orog en ous c e ll . Activity of Primary Parietal cell depen ds on type of plant s. If pla nt belongs to gam opetal ae the n it forms tenuinuc ellate type ovule an d if plant belongs to polypetalae then it forms cr a ssin u cel l ate type of ovule. The prima ry sporogenous cell dire ctly a ct as a me gas pore mother c ell . It divides m ei o t ic a l ly to for m, fo ur ha ploi d m eg as p ores .
The four haploid megaspores generally arranged in linear tetrad. Generally the lower most or chalazal me gas po re remai ns fun ctiona l out of tetrad of me gaspore s and t he ot he r thre e lie towards the mi cropyle degenerate.
This functional megaspore produces female gametophyte. In mos t of Angiosperms [C apsella] , Chalazal megaspore remains functional.
[D] DEVELOPMENT OF EMBRYOSAC OR FEMALE GAMETOPHYTE–
Meg agam etog enes is : - M e g as p ore is the first cell of th e fe male gam etophyt e . T his m egasp ore grows in size and obtains nutrition from the nuc ellu s . The nucleus of megaspore divides mitotically to form a two nuclei.
Each nucleus moves towards the opposite pole and reached at their respective poles. Both the nuclei lie at poles divide twice mitotically. Resulting, four-four nuclei are formed at each poles [ Total 8-nuclei].
Out of the four, one-one nucleus migrates from the both poles [ one nucleus from chalazal side and one nucleus from micropylar side] towards the centre. They are known as pola r nucle i. Both polar nuclei are present in the centre.
Remaining three-three nuclei at each pole surrounded by cytoplasm to form cells as a result of cytokinesis.
Three cells are formed towards the micropyle in which one cell is large and more distinct out of three cells.
This is called egg cell and remaining two smaller cells are known as synergids . These three micropylar cells collectively known as egg -apparatus . [1 Egg cell + 2 Synergids] The three cells are formed toward the Chalaza are called antipodal cells . Both the polar nuclei present in the central cell. But just before the process of fertilization they unite or fuse together in the centre to form secondary nucleus . It is diploid in nautre [2n] a nd one i n number.
Therefore, sev en cel ls and e ig h t n u c lea ted structure is formed. This eig ht nu cleated and sev en cel led s truc ture is called female gametophyte or embryosac of Angiosperms. This type of embryosac is known as "polygonu m type " becaus e it is discove red by S tra s b urg e r in Polygonum plant. Polygonum type embryosac is most c om mon type in Angiosperms [Capsell a] . Polyg onum type of em bryosa c de ve lops from s i ng le megas pore so it is a lso know n as monosporic embr y osac .
-
Fingers like processes are produced from the outer wall of the synergids are known as fil iform app aratus . With the help of these structures, synergids absorb food from the nucellus and transfer to the embryosac. Filiform apparatus is less developed in antipodal cells. Filiform also secrete chemicals which attracts the pollen tube.
-
In some plants, barrier are present either above or below the female gametophyte . These barrier are made up of thick walled cells of nucellus. They prevent the movement of embryosac towards the chalaza or micropyle.
The barrier which is present towards the c hala za is called "hypostase" e.g. Umb ellife rae family and Zo s te ra a nd Crozophora plan t s.
The ba rrier which is present toward the micr opy le is c alled " ep is tas e" e.g. C os tal ia a nd C os t us .
[ E ] TYPE OF EMBRY OSACS :-
( i ) M onos poric Embryos ac – It is of tw o typ es –
1. Polygonum type – It is eight nucleated and seven celled embryosac.
2. Oen othe r a ty pe – Exception ally it is four nuc leated in whic h only one nucleus in a cent ral cell and three nucleus in egg apparatus. Antipodal cells are absent. Endosperm will become diploid. [Micropylar megaspore become functional]
(ii) Bisporic Embryosac – It is formed by two megaspores. It means it develop from two nucleated megaspores. It is of tw o ty pe s –
1. Allium type – Eight nucleated and seven celled [Chalazal megaspores]
2. Endymion type – Eight nucleated and seven celled [Micropylar megaspores]
(iii) Te trasporic Embr yos ac – It is formed by al l four meg aspore nuclei because meiosis is not accompanied by Cytokinesis, so that four nuclei of megaspores are formed.
All four nuclei are colletively known as "Coenomegaspore".
Generally all the four Nuclei of megaspores take part in the formation of the seven celled and eight nucleated embryosac. Beside this different other types of embryosacs are also formed. Arrangement of nuclei are not definite in embryo sac in some of Angiosperms. So that different type of embryosacs are formed as follows :-
|
Type s of Tetrasporic Embr yosac |
Number of nuclei in Sec.nucleus |
Ploidy of endosperm |
|
1. Ado xa ty pe 2. C hry santhe m um ty pe 3. Plumbago type 4. Plu mba g el la ty pe 5. Fritillar i a type 6. Pe na ea type 7. Peperomia type |
2 3 4 4 4 4 8 |
3n 4n 5n 5n 5n 5n 9n |
=====================================================================
Pollination - Sexual Reproduction in Flowering Plants, Biology, Class 12
POLLINATION
"Pollination is defined as the process of transfer of pollen grains from anther to the stigma of the same flower or of different flower of the same species."
Pollination is of two types :-
1 . SELF POLLINATION OR AUTOGAMY :
If the pollen grain are transferred from an anther to the stigma of the same flower , or different flowers of the same plant is called self pollination or autogamy .
2 . CROSS POLLINATION OR ALLOGAM Y :
When the pollen grains are transferred to the stigma of other flower of the same species is called Cross pollination or Allogamy. It takes place in between two different flowers.
Cross pollination is of two types :-
( i ) Geitonogamy : When, pollination takes place in between the two flowers of the same plant then it is called geitonogamy . From the genetical point of view geitonogamy is self pollination because all flowers of the same plant are genetically identical. But ecologically , it is considered as cross pollination .
(i i) Xenogamy : When the pollination takes place in between the two different flowers of two different plants of the same species then it is called xenogamy . This is real or true cross pollination . Genetically , as well as ecologically , it is cross pollination .

Contrivances or Adaptation for Self Pollination :
(i) Monocliny (Bisexuality) - It means flowers are bisexual.
(ii) Homogamy : - When both the sex organs of a flower mature at the same time. It is called homogamy . It increases chances for self pollination. E.g. Mirabilis , Catharanthus .
(iii) Cleistogamy :- In some plants bisexual flower are formed which never opens throughout the life. Such flowers are called cleistogamous flowers , such as Commelina, Viola , Oxalis, Juncus, Drosera. Commelina plant have two types of flowers. One type of flowers are cleistogamous and anoth er are chasmogamous flowers .
-
A special type of cleistogamy is found in flowers of legume plant. The sex organs are closed in a structure which is formed by joining of some petals . It is called keel . This keel never opens . So only self pollination takes place in these plants.
(iv) Bud pollination:- This pollination occurs in bud stage before the opening of flowers. E.g. Pisum, Wheat, Rice.
Contrivances for Cross Pollination
(i) Dicliny (Unisexuality) :- Presence of unisexual flowers confirm cross-pollination. Self pollination never takes place in these flowers. It means allogamy becomes compulsory. Examples – Morus , Palms, Date Palms, Cucumber and Cucurbita, Carica .
(ii) Dichogamy :- In many bisexual flowers of the plants, stamens and carpels of a flower do not mature at the same time. Dichogamy is of two types –
(a) Protandry :- The anther of a flower mature earlier than carpels, is called protandry . Many plants of Angiosperms are cross pollinated only because of protandrus condition . E.g . Salvia, Sun flower, Cotton.
(b) Protogyny :- The carpels of the flower mature earlier than stamens. It occurs in few plants e.g. Ficus bengalensis , Saraca indica , Ficusreligiosa , Aristolochia, most of plants of Crucifrae and Rosaceae family .
(iii) Chasmogamy or Anthesis :- Opening [blooming] of the floral bud in the form of a flower is called anthesis .
(iv) Herkogamy :- In some plants, morphological barriers are formed in between the anther and stigma of the same flower, so self pollination can not occurs and pollen grains from the anther are unable to reach the stigma of the same flower. In this condition, only cross pollination is possible. e.g. Gloriosa, Calotropis and caryophyllaceae family.
(v) Heterostyly :- There is difference in between the length of the filaments of stamens and length of style in flowers of some plants. Some of the plants having long stamen and short style, and in some of the plants bears long style and short filam ent. Due to this reason, self pollination can not possible in these plants e.g. Primrose, Linum, Primula .
(vi) Self sterility or self incompatibility or intraspecific incompatibility :- In this condition the pollen grains of the flower can not germinate s on the stigma of the same flower. T his condition is called self sterility . This is a parental [Genetical] characteristic feature which is controlled by genes . Such as in Pitunia, Malva, Thea, Passiflora, Vitis, Apple (Malus) .
Incompatibility involves many complex mechanisms associated with interactions of pollen and stigmatic tissues. If the incompatibility is due to the genotype of the sporophytic /stigmatic tiss ues, it is termed sporophytic incompatibility on the other hand, if it is due to the genotype of the pollen, it is termed gametophytic in compatibility . This may be due to prevention of pollen germination, retardation of growth, deorientation of pollen tube or even failure of nuclear fusion . It is controlled by genes with multiple alleles (s-allele). Enlarged pollen tube turns upwards and degenerates in style.
(vii) Prepotency:– Growth of its pollen tube of self pollinated pollen grain is very slow and growth of the pollen tube of cross pollinated pollen grain is very fast so pollen tube of cross pollinated pollen grains reach earlier inside the ovule. This is termed prepotency . e.g, Apple, Grapes
METHODS OF CROSS POLLINATION
1 . ANEMOPHILY :
When the pollen grains are transfer from one flower to the another flower through the air is called a ne moph il y an d flowe r i s known as an emo ph ilous flow ers. Such as Ce real plants :- Maize etc.
Anemophil y is also found in all Gymnos perms . T he an emop hil ous pla nts produce en ormous a moun t of pollen grains . The pollen grains are very s ma ll , li gh t weig ht and dry and t heir stigm a is hai ry or bru shy and muci lagenous (Stic ky) .
Yellow c louds are formed i n the sky during the w i nd pollination in Pinus .
These yellow clouds are formed due to the pollen grains are called " Su lph ur Showers".
Anemophilous flowers are neither attractive nor with fra g ran c e . T hey do not have nec tar g lands .
Anemophilous flowers are generally unisexual.

E .g . - Sug ar cane, B am b oo , Coconut, Canna bis, Gras ses, Date palms , T ypha, Oak, Haze l, Cucumber, Papaya, Cotton, Tobacco,Eucalyptus.
-
P ot a m og e t on a nd Myr iophyllum are aquatic plant but it is anemophilous.
-
Maximum loss of pollen grains takes place only in this pollination. It is completely non dire ction al proc ess .
2 . HYDROPHILY :
When the pollination brings by water is known as hydrophily . It is of t wo types.
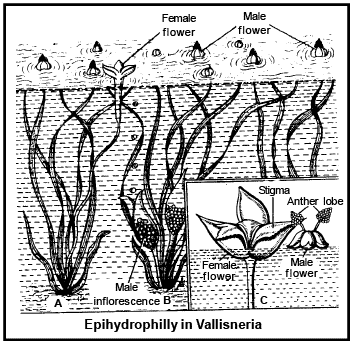
(i) Epihydrophily :- W hen the pollination takes place on the surface of water is called epi hydrophi ly e.g.
Vallis neria and Ru ppi a .
(ii) Hy po hy droph il y : - W he n t he pollination takes– place inside the water is called hyp ohydrophi ly . e .g.
Ceratophyllum, Najas Zo s te ra and H y dr illa .
Spe cifi c g ravit y is found in pollen grains so they remains suspended in water.
3 . ZOOPH ILY :
When the polli nati on brings by animals is called zoophi ly . Generally in zoophillous plants, flowers are very l arg e, at tr ac ti ve an d ma ximum n umbe r of ne ct ar g l an ds are pres ent (more tha n e ntomophill ous ).
(i) ENTOMOP HILY : The pollination takes place with the help of in sec ts is known as en tomophil y. Most of insec t pollination (80%) only by Hon ey be es . Mo st of entomo philou s plan ts are ornam ental plants . Orn am en tal p lan ts utiliz e th eir maximum energy in this pollination and develops different types of adaptation for attraction of insects. These flowers are at tractive i n colour. They posse s s peci a l frag ren ce . Nec tar gl a nd s i s a ls o p re s ent . e .g. Ros e, Lemo n. The pollen g rain s of i ns ec t pollin ated f lowers becom e s tic ky due to pre se nce of pollen kitt . T he surface of stigma of flowers is ro ugh .
Some of the plants develop special adaptation for insect pollination.
-
In Mus saen da pl ant, a dv ert is in g f la g is present for attraction of insects.
-
The lever m echa nism or turn pi pe mechan ism is foun d in Salv ia for poll ination.

The flow ers o f S al vi a have b il ab i at eco ro llat e tube. The connective of stamen is long. The anterior anther lobe of connective is fertile while posteriorly lobe is sterile. When the insect lands on the lower lip, the fertile lobe automatically brings down to touch the back of insect and thus depositing the pollen grains on the back of insect.
-
Some of the flowers have attractive br a c ts i.e. bright and coloured e.g. Boug ain villea
-
Yu c ca pla nt has de velops a n ob lig a te s ym b iot ic relationship with an insect- Pronuba m o th . T he p ol linatio n i n "Yucca take s place only by Pronuba moth. This insect lays eggs in the ovary of flower. Life c ycle of bot h dep end on each other.


-
"Trap door mecha nism " is found i n species of Fic us (Peepal) for pol li nat ion [Blastophaga i nsect] because Hypanthodium type of inflorescence is present.
-
In A ris t o loc h ia "Fly tra p mec hanis m " is fo und for pollinat ion. This flower is known as " Pi t fa ll f low e r" .
-
Ophr ys spec ulum f lower pol li nated by Wasp [Colpa aurea] by mea ns of ps eudo-copulat ion . T he appearence and odour of the flower like female wasp [Mimicry].
-
In Centau rea plant pollina tion takes place by "piston mechanism ".
-
In R af fles ia , the p ol lina ti on is b ro ugh t ab out by Ca rrion fl ie s a nd di sp ersa l o f seed s by El ep han t .
-
Nymphaea , Nelum bo a nd Alis ma are also en tomop hilou s pla n ts w h i le th e y a re hydro p hytes .
(ii) Orn ithophily :-Then process of pollination is taken place by birds is known as ornithophily. e.g. Sun bird or humm ing b ird in Bi gno nia plant and by Honey b ird in S treli tzia , Calli s t em o n ( Bo ttle br us h) , B omb a x [ S ilk c ott on tr ee , ] Bu tea monosperma.

(iii) Chiropterophily :-If the pol lin a tion bring s through the b a t s (Pter o p us ) is c alled chi ropterop hily . These flowers are big in size e.g. Anthoce phalus kadamba, Bauhinia , Ki gelia plants, Adansonia, Mus a.
(iv) Myrmec ophily :-Th is pollinat ion brings about by Termi te s a n d An ts . e.g. P ros opi s (kik ar), Ac acia ( Mimo so i de ae family), Mango and some members of Rubiaceae family.
(v) Mal acop hily or Malm acop hily :-This polli na tion brings about by Sn ai ls e.g., Aspid istra lu rida and Chry sa nthem um plants.
-
Erythrin a (C oral t re e) p la nt v is it ed b y C r o ws and Sq uirrels.
-
In Ar is aema or Cobra Pla nt pollination by s nake c alled "O ph ioph il y".
-
Ni gh t flow eri ng p la nts are pollinated by Moths. They are highly s cented.Their flower generally white coloured
-
Orchids are po l l i n a ted b y W asp s/ Sn ails.
-
Favourable c olour of Honey bee s is yellow , but the y are bli nd to red colour.
-
Larger animals such as some primates (lamurs), arboreal (tree-dwelling) rodents, or even reptiles (gecko lizard and garden lizard) have also been reported as pollinators in some species.
FERTILIZATION
Th e fus ion of male game te wit h fe male ga mete i s ca lle d fe rti lization . Fi rst of all, fertili zation was discovered by S tras bu rg er ( 18 84) in Mon ot rapa plant. This process is completed in the fol lowing steps :-
[ A ] GERMI NATI ON OF POLLEN GRAINS :
Af te r pol li nat io n, pollen grain s g er mi na te o n th e st ig ma . Th ey a bs orb mo is tu re a nd s ug ar c on te nt s from stigma and swellup. The intine of pollen grain grows out through the any one germinal pore of exine, in the form of tube like out growth is called pollen tube. One pollen tube develops in Capsella and mo st of Ang ios perms is called mo nos iph on ous condition, but more than one pollen tubes develops in Malvaceae and Cucurbitaceae fam ily. It i s c alle d p ol ysi ph o no us .
-
W hen the pollen tube come s dow n from the st igma into the st yle, first of al l vegetat ive nucleus enter, into t he pollen tube then it is followed by generative cell. The tube nucleus always occupies in terminal position in pollen tube. The vegetative nucleus controls the growth of the pollen tube. Mean while, the generative cell divide mitotic ally to form two male gametes . Both of the male gametes are non motile .
-
Boron and calcium ions (mainly Boron) are essential for the growth of pollen tube and best temperature for growth of pollen tubs is 20–30 0 C . Pollen tu be shows apical g rowth and chemotropic movement.
[ B ] ENTRY OF POLLEN TUBE INTO OVUL E :
Finally, the pollen tube enters in the ovary at that time, ovule becomes mature. Inside the ovary obtur ators guides the passage of pollen tube towards the micropyle. A mature ovule in which embryo sac also matured, has three paths for the entry of pollen tube:-
( i ) POROGAMY :- In this, pollen tube enters into the ovule through the micropyle. It is known as porogamy. It is found in most of Ang ios per m s [C aps e lla ].

(i i) CHALAZOGAMY :- In this method, the pollen tube enter into the ovule through the chalaza. This method is discovered in C as ua ri na by Tre ub [1891] e .g .
Be tu la and Juglans (w alnut).
(iii) MESOGAM Y :- In this method, pollen tube enter into the ovule either through integu ments - Cucur bita or through the fun ic ulu s - P istac ia and Po pulu s .
[ C ] ENTRY OF POLLENTUBE INTO EM BRYOS AC :
Pollen tube can enter into the ovule through the any passage but inside embryosac, it enter only through the egg apparatus. After the entrance inside the ovule, it grows towards the egg apparatus because synergids cells sec rete the chemical (hormones) which attracts the growth of pollen tube. It means poll en tube shows chemotropic movement in ovule. * Any one synergid starts degenerating when the pollen tube comes near egg apparatus. The pollen tube enter into the embryosac through the degenerating synergids. * When tip of the pollen tube enters into the embryosac vegetative nucleus degenerates. The tip of the pollen tube swells and burst [Endosmosis] after reaching inside the embryosac. The pollen tube released all contents including both male gametes inside the degenerating synergids of embryosac. * Tw o d ar k gr anul es appear s in the r egion o f degener atin g synergids. These are k nown as X-bodies. They are two in no. and both X-bodies are formed by the degenerating nucleus of tube cell and synergids.
[ D ] FUSION OF GAMETES :
-
Before or after the entrance of pollen tube into the embryosac, both polar nuclei of the central cell fused together to form a dipl oid nucl eus . It is known as sec o ndary nucleu s or defi nitive nucle us .
-
Out of two, one male g amete fertilized with egg cell and to form a diploid zygote . This fusion is known as s yn g a my . This is true m echa nism of fer t ilization process.
-
The second male gamete fused with di ploi d s e con dary n uc leu s which is formed by the fusion of two polar nuclei. This fusion is known as triple fusion resulting, a triploid (3n) structure is formed. It is called prima ry endosperm nucleus.
-
Fe rtiliz atio n takes place t w ice at a ti me in A ng ios p erm is c alled doub le f ertiliz at io n .
-
Double fertilization was discovered by "N awa sc hi n " in L il iu m a nd Fr iti ll a ria plan ts.
-
Do uble fer til iz atio n a nd tr iple fusion is t he specific or unive rsal characterist ic of Angios per m. T he re are f iv e nu cl ei and t hree g a me tes p arti cip ate in do ub le f er tiliz at io n.
-
A zygote is formed by true fer tilization (s yngamy] develops into embryo. Triploid primary endosperm nucleus is formed by triple fusion develops into the en dos p erm which is us ed as nutrition for growing embryo.
-
All the remaining cells of embryosac like antipodal cells, synergids degenerate excluding zygote and primary endosperm nucleus after the fertilization. At this time, zygote obtains food from degenerating synergids and antipodal cells.
-
The fertilization in which non motile gametes are carried to female gamete through pollen tube is known as "Siphonogamy".
-
Entry of more than one pollen tube into the ovule leading to occurence of super numerary male gametes is called "Polyspermy."
|
|


DEVELOPMENT OF ENDOSPERM
-
First of all endosperm develops from the primary endosperm nucleus after the fertilization which stored food materials. It is utilized by the embryo during the early development then after at the time of seed germination.
Food is present in the form of starch in endosperm . The endo sperm is of three types on the basis of development :-
1 . NUCLEAR ENDOSPERM :
-
This type of endosperm mostly found in Dicotyledon [Polypetalae] . Nuclear endosperm is also present in Capsella . Such type of endosperm develops by free nuclear divisions of nucleus of primary endosperm nucleus, Thus a multinucleated endosperm is formed. Later on cytokinesis takes place, so that multicellular endosperm is formed.
-
This type of endosperm is the most common in Angiosperms.
-
The milky fluid is found in green Coconut is example of nuclear endosperm , which is called liquid syncytium.
-
In Melas toma , cytokinesis never take place so that it is always remains nuclear endosperm .
2 . CELLULAR ENDOSPERM :
-
This type of endosperm is found in Gamopetalae group. During the development, each division of primary endosperm nucleus is followed by cytokinesis. So that endosperm is remains cellular from the biginning.
3 . HELOBIAL ENDOSPERM :
-
During the development of this type of endosperm, first division of primary endosperm nucleus is followed by unequal cytokinesis so that two unequal sized cells are formed (Cell towards the micropyle is large). Now free nuclear divisions takes place in each cells, results it becomes multinucleated. Eventually cytokinesis takes place later on so that it is changed into a cellular endosperm. This type of endosperm is found in Monocots . It is intermediate type of endoperm.
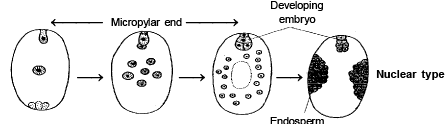
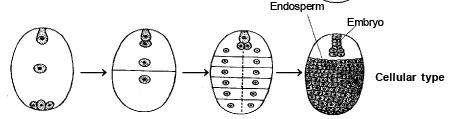
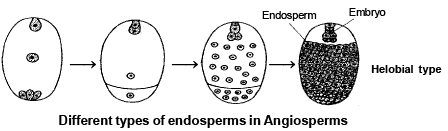
SPECIAL POINTS :
-
After the observation of first two divisions of primary endosperm nucleus, endosperm can be identified whether it would be nuclear , or cellular endosperm .
-
Endosperm is absent in some of Angiosperms e.g. In Orchid aceae , Podostemaceae and Trapaceae .
-
Exceptionally, some of the plants have diploid endosperm instead of triploid such as in Oenothera .
-
Maize and Tom a to have mosaic endosperm in which patches of different colours are present.
-
Theen dosperm in Betalnut is rough surfaced. It is known as "ruminate endosperm " .
-
The drinking portion is nuclear endosperm and edible portion is cellular endosperm in Coconut .
===============================================================
Embryo development - Sexual Reproduction in Flowering Plants, Biology, Class 12
DEVELOPMENT OF EMBRYO IN DICOT
Development of embryo in Capsella is first time discovered by " Hanstein" .
-
In Angiosperm, Zygote undergoes in resting phase. When the endosperm is formed, development of zygote starts. In the beginning it absorb food from the endosperm and increase in size then after a layer secreted by itself. Now it is called Oospore .
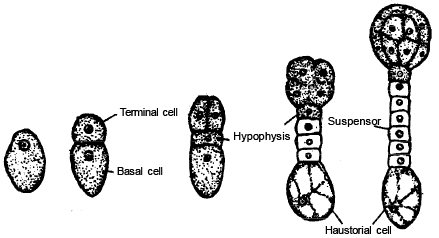
-
The first division of Oospore is transverse , results two cells are formed. The one cell lies towards micropyle is called basal cell or suspensor cell . The other cell is formed towards the Chalaza is called apical cell or terminal cell or embryonal cell .
-
The basal cell and embryonal cell divide simultaneously
-
The basal cell divides transversly and apical cell divides vertically resulting, two suspensor cells and two embryonal cells formed. This stage is made up of four cells which are arranged in ' T' shaped structure. Embryonal cells divide vertically to form four Embryonal cells. This is the quadrant stage of embryo.
-
The two suspensor cells divided by transverse divisions forming a 6-10 celled long filament like structure is termed suspensor. The main function of suspensor is to pushe s the developing embryo into food laden endosperm to provide nutrition.
-
The micropyler cell of the suspensor swells up. This cell of suspensor is known as haustorial-cell .
-
The cell of suspensor lies near the embryonal cells is called hypophysis. T his cell combined with radicle to form the apex of root [Root cap].
-
These four cells quadrent embryo further divide transversely to produce eight. The eight ce lled stage of embryo is called octant stage . The eight cells of octant are arranged in two tiers.
-
The four cells of the octant embryo lies near the hypophysis is known as hypobasal cells and four cells present towards the chalaza termed epibasal cells .
-
The hypobasal cells gives rise to radicle and hypocotyl and epibasal cells giverise to two cotyledons and plumule of the embryo.
-
All the cells of octant divided by periclinal division so that a 16 celled globular embryo is formed [Proembryo].
-
Due to fast division of embryonal cells of globular embryo, a heart shaped embryo is formed. All the cells of this embryo are meristematic.
-
Due to the fast growth in two lobes of heart shaped embryo, they develop into two cotyledons . Both the growing cotyledons turn in downwards due to the curved position of body of ovule of Capsella .
-
The tissues are present above the joining place of both the cotyledons are responsible to form plumule and behind it epicotyl is formed.
-
The tissues present opposite to the plumule give rise to radicle .
-
This curved position of the embryo is called Torpedo or Chordate stage .
-
An axis is present between plumule and radicle is called embryonal axis . It is also called Tigellum [ main embryonal axis].
-
Both the cotyledons are present at lateral position of embryonal axis and plumule is formed in terminal position in Dicotyledon embryo.
-
This type of development of embryo i s known as Cruci fer type or Onagrad type . It is the mos t c om mon ty pe of development in Di c o ts .
-
Crucifer type of development is also found in Capsella, so that it is considered as typical Angiosperm for the s tud y of em br yonic deve lo pm ent of A ngio sper ms .
-
During the development of embryo, embryo is formed from the some part of zygote so that is called meroblastic development.
-
Ovule m odifi ed in to s e ed in which t es ta i s f orm ed by o uter i nteg um en t and teg m en i s fo rme d by i nn er integument.
-
Only mi c ropy le of ovu le remains unc hange d and also present in s e ed .
-
Entire o va ry modified into fruit . This fruit is formed by fertilized ovary so that it is called true fruit .
-
In some of the Angiosperms fruit is formed from the ovary without fertilization known as parthenocarpic fruit.
-
In some fruit parthenocarpy is useless (If edible part is endosperm or seed)
|
STAGES IN THE DEVELOPMENT OF EMBRYO IN A DICOT |
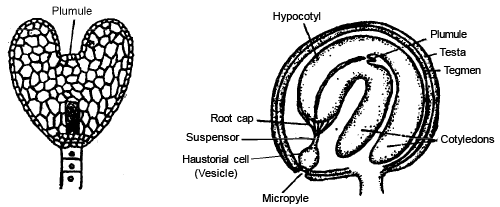
DEVELOPMENT OF EMBRYO IN MONOCOTYLEDON
The Lil ium ty pe of emb ryonic development is found in monocotyledons. The first division is transverse division in Oospore. Results two cells are formed the upper cell chalazal i s called emb ryona l cell and lower micropylar cell is termed as basal cell. The basal cell does not divide further and later on it increases in size and form single celled ve si cul ar suspensor .
Only embryonal cell divides transversly in which terminal cell is called c o ty ledon cell and lower (Middle) cell is known as embryonal axis cell.
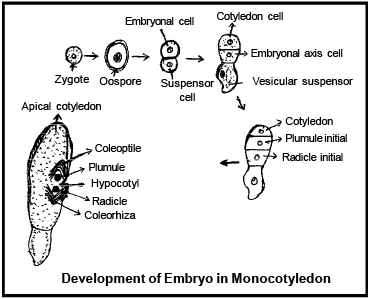
A transverse division takes place in embryonal axis cell to gives rise two cells.
The one cell out of two, gives rise to plumule initial and another gives ris e to radicle initial .
-
The plumule initial divides to form a plumule of the embryo.
-
Radicle initial divides to form radicle. In this both the initials are responsible to form embryo in Lateral position.
-
An apic al c otyled on is formed by the continuous divisions of c otyled on c ell .
-
In Angiosperms, dev elop ment of embryo is meroblast ic a nd endoscopic [towards the chalaza].
-
In the grass family the cotyledon is called scutellum that is situated towards one side (lateral) of the embryonal axis. At its lower end, the embryonal axis has the radical and root cap enclosed in an undifferentiated sheath called c oleo rrhiza .
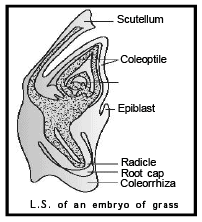
The portion of the embryonal axis above the level of attachment of scutellum is the epicotyl. Epicotyl has a shoot apex and a few leaf primordia enclosed in a hollow foliar structure, the coleoptile.
Reproduction in Plant
Development of floral organs in Arabidopsis
Arabidopsis thaliana is a small weed belonging to family Brassicaceae . It contains approximately 26000 genes and the complete gene sequence has been studies by plant biologists . Like other plants in Arabidopsis the initiation of flowering is influenced by various environmental factors, Of these photoperiodism and temperature are more important. Under the influence of the two factors the apical merisem starts producing flowers instead of vegetative structur. The development of calyx, corolla, androecium and gynoecium is controlled by specific genes. These are called homeotic selector genes , e.g. Apetala-2, Apetala-3, Agamous etc. As a result of effect of these genes, the development of leaves is replaced by the develop ment of flowers.
Similar to leaves, the floral organs developed by periclinal divisions in protodermal or subprotodermal cells. Initially these cells show periclinal divisions followed by periclinal as well as anticlinal divisions resulting in formation of an outgrowth. Cells of the outgrowth divide, develop and differentiate to form different floral organs. In Arabidopsis calyx are the first to develop followed by corolla, androecium and gynoecium. The order of development of floral may vary from plant to plant. For example in membes of family Umbelliferae the sequence of development of floral organs is - androecium, corolla, calyx and gynoecium.

Difference between Self and Cross pollination
|
S.No. |
Self pollination |
Cross pollination |
|
1. |
Pollen grains are transferred to stigma of same or genetically similar flower. |
Pollen grains are transferred to stigma of genetically different flower. |
|
2. |
Anthers and stigma mature simultaneously. |
They mature at different time i.e. protandry and protogyny. |
|
3. |
It occurs in open as well as closed flower. |
It occurs in only open flower. |
|
4. |
It is very economical for plants. |
It is not economical as the plant has to produce large number of pollen grains, nectar, scent and colouration . |
|
5. |
External agencies are not required. |
It is essential i.e. depends on agencies. |
|
6. |
Young ones are homozygous. |
Young ones are heterozygous. |
|
7. |
It produces pure lines because of the non occurrence of genetic recombinations. |
It produces variations due to genetic recombinations. |
|
8. |
It cannot eliminate harmful traits. |
It can eleminate harmful traits. |
|
9. |
Useful characters are preserved. |
Not preserved. |
|
10. |
No adaptability in the changing environment. |
It provides adaptability. |
|
11. |
It cannot introduce new traits. |
It can introduce new traits. |
|
12. |
Disease resistance is low. |
Disease resistance is optimum. |
|
13. |
There is decrease in yield. |
It increase the yield. |
|
14. |
It does not help in the development of new species |
Helps in the development of new species. |
Difference between Embryo and Endosperm
|
S.No. |
Embryo |
Endospe rm |
|
1. |
It is formed by fertilized egg (syngamy) |
It is formed by fusion of secondary nucleus (triple fusion) |
|
2. |
It is always diploid structure |
It is g enerally trip loid struc ture |
|
3. |
t gives rise to new plant |
It provides nutrition to the developing embryo. |
|
4. |
Cotyledons, plumule and radic le are formed in embryo |
Such type of structures are never formed. |
|
5. |
Embryo is pres ent in s eed. |
It is only found in endospermic seeds otherwise It degenerates with the formation of seed. |
|
6. |
Embryo dies in the absence of endosperm |
Endosperm does not die in the absence of embryo. |
|
7. |
Germination of seed takes place. |
There is no germination is found in endosperm. |
Difference between Egg cell and Secondary nucleus
|
S.No. |
Egg c ell |
Sec ondary nucleus |
|
1. |
It is present near the micropyle, inside the embryosac. |
It is present in the middle of the embryosac |
|
2. |
Generally egg is surrounded by two synergids. |
It is not surrounded. |
|
3. |
Only single nucleus is present in it. |
It is formed by fusion of two polar nuclei. |
|
4. |
It is haploid struc true. |
It is diploid structure. |
|
5. |
It is fertilized with one male gamete and to form a diploid zygote (Embryo) |
It is fused with one male gamete and to form a triploid primary endosperm nucleus (Endosperm) |
Difference between male and female gametophyte
|
S.No. |
Male Gametophyte |
Female Gametophyte |
|
1. |
It is developed from microspore or pollen grain. |
It is developed form megaspore. |
|
2. |
It does not remain embedded permanently in m icrosp orangi um. |
It remains embed ded perm anen tly in m ega sp orang iu m. |
|
3. |
Male gametes come out of pollen grain due to the formation of pollen tube. |
Female gamete always remains inside, cove red by m embrane of meg asporang ium. |
|
4. |
There are two phases of growth - prepollination and post pollination. |
Only single phase of growth. |
|
5. |
It is three celled structure in mature stage. |
It has seven cells in mature stage. |
|
6. |
It will disintegrate after fertilization. |
Two new structures are formed after fertilization, that is endosperm and oospore. |
Differe nce betwee n Epigeal and Hypogeal germination
|
S.No. |
Epigeal germination |
Hypogeal germ ination |
|
1. |
Cotyledons c ome above surface of soil during the germination and remains attached till the formation of new leaves. |
Cotyledon remains under the soil surface. |
|
2. |
Due to the growth of hypocotyl, plumule and cotyledon pushed above the ground. |
Due to very less growth of hypocotyl, cotyledons do not come above the ground. |
|
3. |
Growth of epicotyl does not take place. |
Due to high growth of epicotyl, plumule comes above ground and cotyledon remains inside the ground. |
|
4. |
Cotyledons bec ome green and perform leaf like func tion |
Cotyledon remains under the soil and do not perform lea f like func tion |
|
5. |
Cotyledon is responsible to form first leaves, than after dries and fall on the ground |
Cotyle don decomposed inside the soil. |
NUMBER OF CHROMOSOMES IN DIFFERENT PARTS OF PLANT
|
S.No. |
SPOROPHYTE |
|
|
1. |
Zyg ote |
2n |
|
2. |
Emb ryo |
2n |
|
3. |
Endosperm |
3n |
|
4. |
Radicle |
2n |
|
5. |
Plum ule |
2n |
|
6. |
Cotyledon |
2n |
|
7. |
Nuc ellus |
2n |
|
8. |
Integument |
2n |
|
9. |
Microspore mother cell |
2n |
|
10. |
Megaspore mother cell |
2n |
|
11. |
Ovary wall |
2n |
|
12. |
C arp el |
2n |
|
13. |
Sepal, Petal |
2n |
|
14. |
Stamen |
2n |
|
15. |
Leaf, Root, Stem |
2n |
GAMETOPHYTE
|
S.No. |
P arts of Plants |
Number of Chromosomes |
|
1. 2. 3. 4. 5. 6. 7. 8. 9. |
M icros pore Tube cell Male gamete Megaspore Embryosac Synergid Antipodals Egg cell Polar nuclei |
n n n n n n n n n |
Differ ence b etwe en pollination and fert iliz ation
|
S.No. |
Pollination |
Fertilization |
|
1. |
ra nsfer of pol len grai ns from anther to stigma of the flower is called pollination. |
Fusion of male gametes and egg cell in embryosac situated inside the ovule is called fertilization. |
|
2. |
This proces s take place before fertilization. |
This process take place after pollination. |
|
3. |
For the completion of this process, ins ects, water, air like agencie s are e ssent ial |
There is no any external medium is utilized in this process |
|
4. |
Pollen tube is not formed |
Pollen tube is formed which transfer male gametes up to egg cell. |
|
5. |
This process take place on outer parts of flower, so that it is external mechanism |
This process take place inside the flower, so that it is internal mechanism. |
Difference between Monocotyledonous and Dicotyledonous seed
|
S.No. |
Mon ocotyledon ous See ds |
Dicotyledonous Seeds |
|
1. |
Only single cotyledon is present with embryo |
Two cotyledon are present with embryo |
|
2. |
Generally cotyledon is thin or papery |
Generally seeds are non endospermic, some tim e s may be endospermic |
|
3. |
Gene rally seeds are en dospermic |
Not called by this name |
|
4. |
Cotyledon is also called scutellum |
Coleoptile and coleorhiza are not formed. |
|
5. |
In seed plumule is covered by coleoptile and radicle is covered by coleorrhiza. |
Plumule is in termin al position and cotyledons |
|
6. |
Plumule is in lateral position and cotyledon are is in terminal position |
Plumule is in termin al position and cotyledons are present in lateral position |
|
7. |
Radi cle de generates af ter so metime and adventitious roots are formed at that place. |
Radicle is responsible to form primary root. |
|
8. |
In some of the seeds, seed coats and cotyledon fused together e.g. Wheat etc. |
Such types of seed are not found. |
|
|




|
|



============================================================
ASEXUAL REPRODUCTION in plants - Sexual Reproduction in Flowering Plants, Biology, Class 12
ASEXUAL REPRODUCTION
In asexual reproduction, the new individual are produced by any means other than the fusion of sex gametes.
It means a "reproduction in which new individuals are formed without meiotic division and fusion of gametes is called asexual reproduction ." In this way asexual reproduction is also known as apomixis . [Greek - Apo = w ithout; mixis = mixi ng] Apomixis term sugge sted by Winkler.
The Apomixis is characterised by quick multiplication and reproduction of genetically similar plants from the single parent. Such a population produced from single individual is called "clone" and each member of the clone is called ramet .
In flowering plants, there are two main types of Apomixis .
(i) Agamospermy
(ii) Vegetative Propagation
(I) AGAMOSPERMY :
In this type of method embryo is formed without fertilization and meiotic division is called agamospermy . It means plants belonging in this category propagated through seeds but the embryo formation does not involve meiosis and syngamy.
There are three different type of agamospermy –
[ A ] DIPLOSPORY :
In this method archesporium differentiates to form a megaspore mother cell but this megaspore mother cell directly gives rise to an embryosac without meiosis. This embryosac is diploid and a diploid embryo is formed without fertilization from diploid egg of this embryosac .
Example : Parthenium, Taraxacum
|
Parthenogenesis : In this process haploid egg cell of female gametophyte is responsible to form a haploid embryo without fertilization. |
[ B ] ADVENTIVE EMBRYONY :
In this method, an embryo is formed from any diploid cells [Nucellus or integuments ] of the ovule except embryosac. This diploid cell behave like a zygote. Adventive embryony derived from Nucellus in Citrus, Mangifera, Opuntia, Mamillaria and from Integuments in Spiranthus australis .
[ C ] APOSPORY :
It is dicovered by Rosenberg in Heiarcium plant. In this method embryosac or female gametophyte is directly formed from any diploid cell of the sporophyte except megaspore mother cell without meiosis is known as apospory . In this gametophyte always remains diploid , e.g. Heiracium, Ranunculus and Rubus .
POLYEMBRYONY
Many embryos are formed inside the single seed is called polyembryony . first of all, it is observed by Leeuwenhoek in Citrus (Orange) seeds . Polyembryony is commonly found in Gymnosperms but it is also found in some of Angiospermic plants such as Orange, Lemon and Nicotiana etc.
If polyembryony develops naturally then it is called spontaneous polyembryony [self] . When it is developed artificially is called induced polyembryony .
When many embryos are formed from separate-separate embryosacs [more than one] inside the ovule is called false or pseudo polyembryony .
When many embryos are formed inside the single embryosac of the seed is called " true poly embryony". It is either develops from :-
(i) Clevage of Zygote or budding - e.g. Cymbidium, Exocarpus, Nymphaea, Nicotiana, Orchids etc.,
(ii) By the fertilization of synergids – e.g ., Anemone , Aristolochia, Sagittaria etc.
(iii) F ertilization of antipodal cells - e.g ., Paspalum , Ulmus etc.
Note : In Lilium all three types of polyembryony are present. [Adventive embryony is also example of polyembryony in which additional number of embryos are formed from nucellus or integuments ]
ANDROGENIC HAPLOID PLANTS
This concept was given by Haberlandt and practically proved by Steward. It is based on totipotency.
Anther of the plant grown on culture medium by Guha and Maheshwari . As a result of this culture, haploid and diploid two different type of plants are formed. Diploid plants developed from the wall of the anther and haploid plants developed from the pollen grains . Such type of haploid plants obtained from the tissue culture are known as androgenic plants .
Fast vegetative multiplication through the tissue culture from any part [cell or tissue] of the plant in the field of Agroforestry and Horti culture is called "micropropagation ".
(i) P.Maheshwari established – Research centre of Angiosperm Embryology at Delhi University.
(ii) Book published by Maheshwari - An Introduction to the Embryology of Angiosperms .
VEGETATIVE REPRODUCTION /PROPAGATION
Plants belonging to this category propagate by a part of their body other than a seed. The structural unit that is employed in place of seed for the propagation of new plants is called propagule . In Angiosperms any parts of the plants - roots, stems and leaves used for veg etative propagation.
Generally method of vegetative propogation have been further divided into two types -
[ A ] NATURAL VEGETATI VE PROPAGATION :
( a ) By roots :- Modified tuberous root of Sweet potato (Ipomoea batatas ), Aspar agus , Tapioca , Yam, Dahlia and Tinospora can be prop agated vegetatively when planted in soil.
In some plants adventitious buds develop on the ordinary roots like-Dalbergiasisso, Populus , Guava, Murraya , Albizzialebbek etc. which grow to form new plants.
The buds present on the roots grow into leafy shoot above the ground is called Slips.
( b ) Under ground Stem :- In some plants under ground modified stem such as -Rh izomes - Typha; Ca nna, Gi nger , Turmeric, L otus, Musa etc.
Corm - Glad iolus, Col o casi a, Cr o cus, Am o rpho phall us, Alocasi a etc.
B ul bs - O ni on, Ga rlic and L il ie s Tubers - Po ta t o, H el ia nt hu s t ub eros us e tc . wh ic h gro w to form a n e w p la nt s.
Sucker - M int and Chrysa nthemum ;
( c ) Creepers : In creeping stem of the plants adventitious root are developed from the nodes and to form a aerial shoots such as (i) Ru nne rs - Cy no don , Oxa li s and Ce nt el la (ii) Stolon = Fraga ria (Strawber ry) & Valli sneria (iii) Offset - Pistia, Eichhornia (water hyac inth) etc Aerial stem - Opuntia
( d ) Leaves : Some plants produce adventitious buds on their leaves e.g., Br yoph yllu m , Be goni a, Strepto carp us, S aint pauli a . These buds remains dormant, when the leaves attached with plants but after separation, when it comes in contact with moist soil develop new plantlets [buds] which form new plants.
-
In Kal a nch oe pla nt, whol e portio n of le af b lade re generate a ne w pl an t.
-
In s ome of t he plant s, fl es hy ax illa ry b uds deve lop from ax is of l eave s are called Bu lbi ls Example – Dios corea, Oxali s, Dentaria, Globba, Agave, Lilium.
-
T here are special t ype of f le shy buds de velop in Aquat ic pla nt are called Turi o ns . e.g. Potam ogeton, Utricula ria.
[ B ] ARTIFICIAL PROPAGAT ION :
Gardeners and Horticulturist have employed the various method of vegetative propagation for economic production . All the methods a re man made so that their practices co nstitute artific ia l mean s of v egetative propagation. These are as follows
[ i ] Cutting :- A cutting is separated portion of root, stem or leaf which is used for propagation, Factors are taken into consideration for successful propagation such as
(i) age of the parent plant,
(ii) length and diameter of the cutting,
(iii) season and
(iv) type of plants.
Some time the stem cuttings are treated with rooti ng hormone [IBA, IA A or NAA] for prope r develop ment of ad ventitiou s roots e. g., Sug ar ca ne, Rose, Croton, Tapioc a, C hina rose (Stem cut ting) L emon, Tamarind (Root Cut t ing ), Sa ns ev iria (L ea f cu t ti ng ) . [F av oura bl e ti me f or c utti ng - Rai ny s ea so n]
[ ii ] Grafting : - It is the most common method of vegetative propogation described by ancient gardeners long before the science of horticulture became established. In this method, parts of two plants are joined in such a way that they grow as one plant. Grafting is done between two closely related di cot yled onous plants having vasc ular camb ium . The rooted s upported po rtion of one plan t c alled Sto ck is joined with a twig of another plant called Sc ion . Generally, the root s tock belong to wild variety which is resist ant to disease & pe st . The sci o n is derived from the plant possessing be t te r c h ara c ters . In grafting, the stock and scion cuts are marked in such a oblique manner so that they fix with each other.
This joint covered with clay or a layer of wax. Within a few days, tissues of stock and scion combined togehr and to f orm a new plant. e.g . , Gr afted Mango, Roses, o r a nge, Seed less Gr ape s and Guava , Apple , Pear. [Favourable t ime for graf t i ng - Spring se ason]
Types of Grafting -(a) To ngue / Whi pgrafting (b) Wedgegrafting (c) Crown grafting (d) Side grafting (e) Approachg rafting
-
Bud g rafting - This method is similar to grafting except that scion in this case consist of a bud along with some portion of bark having intact cambium.
Generally b ud grafting is done d uring the rainy seas on e.g., Rose s, P each .
[ iii ] Layering : - Thi s m eth od is use d i n t ho se pla nts wh ich are h avi ng flexible long branches. In this method, roots are artificially induced on the stem branches before they are detached from the parent plants for propagation. There are two common types of layerings .
( a ) Mound/G round Layering - This me thod is only utilized in herbacious plants. In this method, the lower branch of stem is bent down and partially defoliated and injury is made on defoliated part. It is covered by thin layer of moist soil in such a way that the growing tip of the branch remains above the soil surface. After a few days the pegged portion develops adventitious roots. Then rooted branch cut and separated from parent and grown into a new plant e.g., Jasm in e,strawberry , Raspberry etc.
( b ) Air Layering or Gootee - T hi s me tho d is c omm on ly e mpl oy ed i n c as e of sh rubs a nd trees which do not possess branches near the ground. In this method a ring of bark is removed [girdled] from the aerial branch . This girdled portion is covered with moist grafting clay [ 2parts clay + 1 part cow dung + some pieces of hay + cotton + water] and wrapped with a polythene sheet.
This wrapped portion of the branch is called gootee. Inside the gootee roots developed within a period of a or two month(s) e.g., Litchi , Pome gra nate, O rang e, Lem on, Bo ug ain vil le a, Guava etc.
According to P. Maheshwari :
|
( a ) Merits of Vegetat iv e p ro pagat ion : |
|
( b ) Demerits of Ve getat ive propagat ion : (i) Diseased parents always give diseasd clone. |
|
SIGNIFICANCE OF VEGETATIVE PROPAGATION 1 . It is the only method of reproduction in those plants which have lost their capacity to produce seeds -e.g. Ban ana, Seed l ess gr apes , Orange etc. 2 . Th e plant s w h ic h pro duce sm all q ua ntiti es of s ee ds, p oor v ia bilit y of see d or p r o long ed s ee d dormancy, reproduce only by this method because it is more rapid, easier and less expensive. 3 . Production of these plants more as compared to seeded plants. 4 . The greatest advantage of this method is that a biotype of plant [Original plant] can be retained and multiplied indefinitely without any change or variation. |
DIFFERENCE BETWEEN ASEXUAL AND SEXUAL REPRODUCTION
|
Ase xual R eproduction |
Sexual Reproduction |
|
1 . It does not involve the formation and fusion of gamet es. 2 . New individual develops from one cell or a part of one parent. 3. New individuals are genetically similar to the pare nts. 4 . It involves only mitotic divisions. 5 . It does not require the formation of sex organs. 6 . It does not introduce variability. Hence, has no evolutionary importanc e. |
It involves the formation and fus ion of two gametes, generally distinguished into male and female. New individual develops from zygote or fus ion product of two gametes , whic h may or may not be produced by two parents. New individuals or offsprings are generally different from either of the two parents. It involves meiosis at one or the other stage. Formation of sex organs is a pre-requisite for sexual reproduction. It introduce s variability and is , hence of evolutionary importance. |



|
|

|
|

|
|





===============================================================
Seed - Sexual Reproduction in Flowering Plants, Biology, Class 12
ANGIOSPERMIC SEED
I. STRUCTURE OF SEED
The whole plant exists in a seed. Plant is the main base of whole living world . So it can be say that seed is the most important structure. In angiosperm, after the fertilization , egg cell changed into embryo, ovary changed into fruit and ovule change into seed .
Morphologically, ripened ovule is known as seed . In other words, seed is amature, integumented megasporangium.
Seeds are characteristic of Spermatophytes (Gymnosperms and Angiosperms). Typical mature seed is having three main parts :
1. Seed coat : Outer, protective covering of the seed is called seed coat, which develops from integuments of ovule. In seeds developing from bitegmic ovules, there are two distinct layers in seed coat. The outer layer is thick, hard and leathery (developing from outer integument), called testa , whereas inner layer is thin and papery or membranous (developing from inner integument), called tegmen . In seeds developing from unitegmic ovules there is single layered seed coat . The seed coat performs usual protective function.
The seed is attached to the fruit wall or pericarp by means of a stalk called funicle or funiculus . The point of attachment of funiculus to the body of mature seed is called hilum. A small opening or pore called micropyle is present just near the hilum, which is the way of entry of water into the seed. As most of the ovules in angiosperms are inverted or anatropous, so a ridge called raphe is present.
2. Embryo : Embryo is the most important part of the seed , which represents minior tiny future plant. Embryo develops from fertilized egg [zygote].
The embryo is having an embryonal axis or main axis called tigellum, to which one or two cotyledons (seed leaves) are attached, depending upon whether the seed is monocot or dicot . The portion of embryonal axis or tigellum below the point of attachement of cotyledons, is called hypocotyl, which bears radicle or future root at its tip. Similarly, portion of embryonal axis or tigellum above the point of attachment of cotyledons, is called epicotyl, which bears plumule (future shoot) at its tip.
In some seeds (e.g. legumes), reserve food is s tored in cotyledons , whereas in others (e.g . cereals), there is special nutritive tissue called endosperm. The seeds having endosperm are called endospermic or albuminous seed, e.g ., Cereals, Castor , etc . , whereas seeds in which endosperm is fully consumed by embryo and noendoserm is left, are called non-endospermic or ex -albuminous seeds , e .g., Gram , Pea, Sem, Cucumber , Tamarind , etc. here serve food materials in seeds may be carbohydrate (e .g., Wheat, Rice ) or proteins ( legumes) or fats ( Castor, Peanut, Sunflower ), etc.
Some time, some part of nucellus remains unused which is present in the form of thin layer around the endosperm is called perisperm . e.g. Betelnut , Black pepper, Castor .
-
In Monocotembryo , there is a single cotyledon called scutellum. e.g. Grasses like Wheat. [On opposite side of scutellum is a tongue shaped outgrowth remains of second cotyledon is present called epiblast. ]
-
In Dicot albuminous seed-castor (Ricinus communis), there is a specific outgrowth called caruncle or strophiole, present over micropyle. It is formed by proliferation of cells of outer integument at tip. Caruncle is somewhat spongy and helps in absorption of water during germination of seed.
-
On the basis of cotyledon in the seed of angiosperm are called Monocotyledon [one cotyledon] and dicotytedons [Two cotyledon ].
3. Endosperm : It is the nutritive tissue which may present or absent in the seeds. The angiospermic seed classified into two categories on the basis of presence or absence of endosperm in seeds -
1. Non Endospermic or Ex-albuminous seed
2. Endospermic or Album inous seed.
1. Non Endospermic or Ex-album inous seed : Such type of seeds do not have an endosperm at maturity, therefore are called non endospermic or Exalbuminous seeds. The endospermic tissues are absorbed during the development of embryo. The absorbed food materials from the endosperm is stored in cotyledons, that why they become so large and fleshy e.g.
Capsella and most of dicotyledons. But Castor seed is endospermic.
Non-endospermic dicot seeds : e.g., Gram , Bean, Pea, cucumber ,Tamarind.
Non-endospermic monocot seeds : Pothos (money plant), Vallisneria , Alisma , Amorphophallus .
2. Endospermic or Albuminous seed : This type of seeds, food is stored in endosperm . Such seeds are called endospermic seed OR Albuminous seed. The endospermic tissue in these seeds utilize during the germination of seed and their cotyledons are thin and membranous e.g. Most of Monocot seeds e.g. Wheat, Rice , and Maize etc.
Endospermic dicot seeds : e.g., Castor, Papaya, Cotton.
Endospermic monocot seeds : e.g., Maize, Rice, Wheat and Coconut.
II. STRUCTURE OF BEAN SEED :
Bean seed is kidney shaped having a concave side and a convex side. The concave side is slightly darker and posses a white elongated scar is called hilum, which marks the point of attachment of the seed with the funiculus. A small pore lies adjacent to the hilum, called m ic rop y le, which performs the function of absorbing water during germination. There is a short ridge on the other side of hilum. It is called ra phe.
The Bean seed has two see d c o ats . The outer seed coat is called test a. It is hard, thick, s mooth and variously coloured. It develops from outer integument of ovule. The inner coat is white, membranous and thin. It is called tegmen. It is difficult to separate tegmen from testa. The seed coats can easily be removed if the seeds are soaked in water for a few hours.
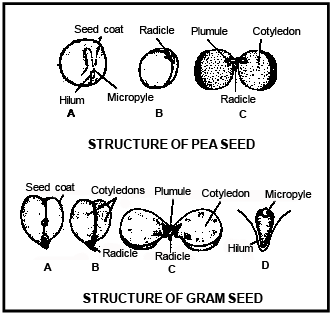
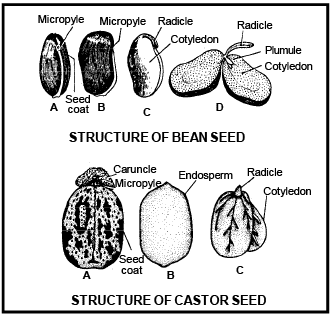
Each seed encloses an embryo which can be examined after removing the seed coats. The bulk of embryo consists of two large, white and kidney-shaped cotyledons (or seed leaves). They store re serv e food material in the form of carbohydrates and protei ns to provide nourishment to embryo at the time of seed germination and growth of young seedling. The cotyledons are attached laterally to the curved embryonal axis at the cotyledonary node. The micropylar end of embryonal axis is rod-shaped and slightly bulged. It is called radicle (or the embryonic root). The other side of embryonal axis lies in between the two cotyledons. It is called plumule (or the shoot tip). The shoot tip remains protected by two small, folded leaves. The portion of embryo axis between the plumule and cotyledonary node is called epi coty l. The portion of axis between the radicle and cotyledonary node is c alled hyp ocotyl.
STRUCTURE OF MAIZE GRAIN [SEED]
A grain of Ma ize (Z ea m ays ) is infect a dry one-seeded indehisc ent frui t of c ary ops is type in which pe ri ca rp (or fruit wall) is fused with the seed coat. The covering of grain enc loses tw o ma jor p art s-en dos pe rm and an embryo. As seen in longitudinal section, the major portion of grain is occupied by a large starchy e ndosperm.
It i s sur ro unded by a sp ecial one-cel l ed t hick layer, called al euro n e l ay er. T he cel ls o f ale urone l ayer are filled with proteineous grains which play an important role at the time of germination.
The embryo consists of an upper shield-shaped sc u tellum which is actually a cotyledon. The scutellum is closely pressed against the endosperm and helps in the translocation of nutrients from endosperm to the growing embryo at the time of germination and seed growth. The epidermal tissue of scutellum lying in direct contact with the endosperm is both secretory and absorptive in function. It secretes certain hormones into the endosperm which synthesize enzymes. These enzymes are responsible for the decomposition of stored nutrients and convert them into soluble state. The soluble nutrients are then absorbed by ep ithe lial l ayer of scutellum and transferred to the g row i ng em b ryo .
The scutellum is attached laterally to the embryonal axis. One end of the embryonal axis points towards the p oi nt ed e nd of the grain. It is called rad ic le. The radicle is covered by a protective sheath called coleorhiza.
The other end of embryonal axis, which faces towards the broader end of the grain, is called plumule. The plumule is s urrounded by a conical protective shea th, called coleoptile. The coleoptile is capable in growth and covers the growing shoot tip till it passes through the soil during germination.
|
A STRUCTURE OF MAIZE SEED |
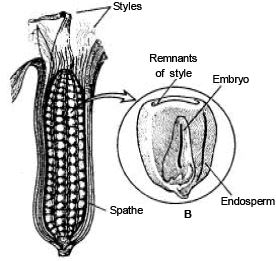
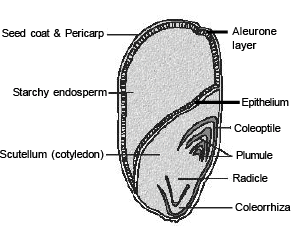
II. G ER MI N AT ION O F S EE D :
The process by which the dormant embryo wakes up [becomes active] and begins to grow is known as germination of seed. T he embr yo of seed grows in to a spo rophyte plan t. The radicle produces primary root. The plumule develops short and latter on cotyledons degenerates.
Types of seed Germination : Germination is basically of two types, depending upon behaviour of cotyledons but viviparous germination also found :-
1. Epigeal germination : Here due to hypocotyl growth or elongation, cotyledons are pushed out of soil. This is called epigeal germination.
T his t ype of germi nat ion occurs i n Co t ton , Pa paya , Ca stor, On ion, Cuc urbi t s, Tama ri nd , French bea n, mu s t ard , etc. In some c ases, thes e above ground c ot yle don s beco me gre en lea f like (cotyledonary lea ves) and perform photosynthetic function till the seedling assumes independency (e.g., castor, cotton, onion, papaya, etc.). In ot her plants, cot yledons do not assume leaf like shape a nd fall off (e.g., Fren ch b ea n a nd Tamarind).
2. Hypogeal germination : Here due to growth in epicotyl plumule comes out of the ground and cotyledons remain underground. This is called hypogeal germination. This type of germination occurs in most of the monocytyledons and few dicotyledons, e.g., Maize, Rice, Wheat, Coconut, Gram, Pea, Pe anut and Mango , etc.
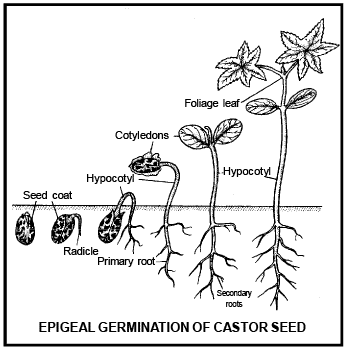
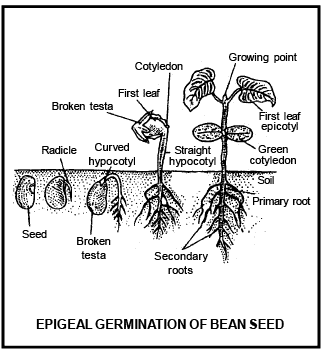
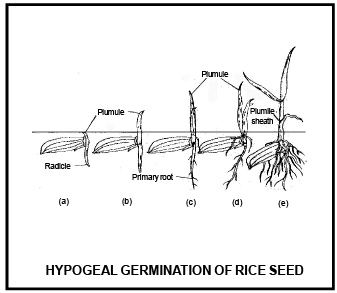
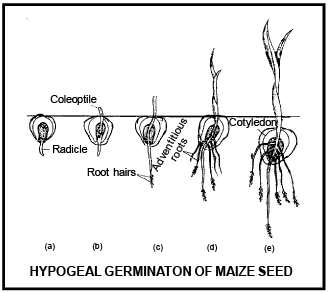
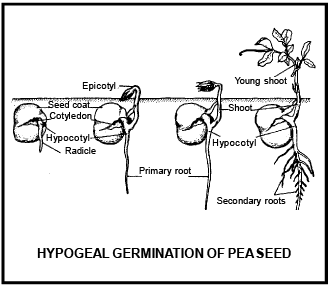

3. Vivipary : It is a special type of seed g ermination which is characteristic of Mangrove vegetation, found in muddy, saline conditions , e.g., Rhizophora, Avicennia, Ceriops, Breguira, Sonneratia, etc. Here there is no resting period of embryo and germination occurs inside the fruit, while it is attached to the parent plant, i.e., "in-situ germination".
The seedling is separated in the mud with the help of lateral roots developing from basal end of radicle. This is called viviparous g ermination or vivip ary.
III.. FACTORS AFFECTING SEED GERMINATION :
The seed germination takes place in the presence of favourable condition such as temperature , moisture , and air . This is a very delicated stage in the life of the plant. Any minute change in the environment have harmful effect on growing embryo. So favourable environment should be essential for germination. The following factors are essential for germination of seed.
1. Moisture :
The moisture or water is the most important factor for germination of seed. Generally, the cells of embryo contain about 10 -1 5% water in the dormancy period. The vital activities like growth and development is unable to continue in this less amount of water. For active life processes, water must be pre sent about 75-90% . The seed absorbs water and swell up to increase in size before the germination. Water absorbs through seed coat and micropyle . The following effects have been observed in seed due to absorption of water -
I . The seed coat becomes soft and absorbed water reaches to the cells of embryo. When the ratio of water increases in the cells, the protoplasmic contents become more active.
II . The size of embryo increased due to water absorption due to this, seed coat breaks up and radicle and plumule easily comes out. Seed coat absorbs water, it becomes soft and increase its permeability. Resulting it increases rate of exchange of oxygen and CO2. The rate of respiration increases in active cells of embryo. Thus it requires more oxygen.
III . The enzymes are present in the cells of endosperm or cotyledons which are active only in the presence of water. The stored food changed into dissolved form by the activity of enzymes and it reaches to the active embryo.
2. Oxygen [O2 ]
The process like cell division, cell elongation etc. of the embryo require energy . This energy is released by the oxidation of organic substances. Oxygen is essential for oxidation process. The upper surface of soil contains sufficient amount of O2. The healthy germination does not take place in the absence or scarcity of oxygen in deep soil so crop seed sown in the soil by the farmers upto 5-7 c.m. deep.
3. Temperature :
The suitable temperature is essential for germination of seed. The protoplasm of the cell remains active at certain range of temperature. Most of seed do not germinate in between the range of 00 C to 50C and above the 45 0 C. The favourable range of temperature is 20-250 C for germination of seed.
4. Food or Nutrition : The growing embryo requires [needs] nutrition during germination. The embryo depends upon stored food materials in cotyledons or endosperm in the germination period upto the formation of primary root from the radicle and first leaf from the plumule.
5. Light :
Light produces different effect in different variety of seeds. Most of the plants do not require light up to the formation of first leaf. The rate of germination very fast in the absence of light. But for some plants light is very essential for germination. They will not germinate in the dark. For example seed of Orchids, Tobacco , Mistletoe etc.
After the development of newly leaves on shoot light becomes very essential factor. The chlorophyll is not formed in the absence of light and this new formed plant [juvenile] will die in the absence of photosynthesis and deficiency of food.
IV. SPECIAL POINTS
1 . Highest amount of fat is found in endosperm of Coconut.
2 . Embryonic development of Capsella is endoscopic because it is developed towards chalazal region of the zygote.
3. Single celled suspensor is found in Triticum [ wheat] and Caryophyllum . Their seeds are caryopsis fruit.
4 . Suspensor is absent in Sunflower and Marigold.
5. 125 meiotic divisions are essential for development of 100 grains of Wheat .
6. Two generation and three genotypic cells are present in Angiospermic seed .
7 . Heaviest seed (6 k.g.) is found in Lodoshia. Its fruit is 1 meter in length and wt. of fruit is 18kg .
8. Edible part of Betelnut is endosperm.
9. Abscisic acid is the natural germination inhibitor of seed.
10 . Anatomical transition from root to stem occurs in hypocotyl region.
11 . Seed of the Ruella dispersed by jaculat or technique.
12. Smallest or minute seeds are found in Orchids which are lightest in plant kingdom and are called " Dust seeds " [w t. 20.33 mg].
13. Xenia (by Focke) - Effect of pollens inside Embryosac on Endosperm (except Embryo) e.g Maize.
14. Metaxenia - Effect of pollen out side the Embryosac on seed coat or pericarp. In Datepalm, maturity time as well as size of fruits can be changed by using different pollens.
15 . The pollen grains of insect pollinated flower bears oily layer is called pollen-kitt. It is made up of lipid and carotenoids.
16. The longest pollen tube is found in Maize .
17. Seeds of a large number of species live for several years. Some seeds can remain alive for hunderds of years. There are several records of very old yet viable seeds. The oldest is that of a lupine, Lupinus arctic us excavated from Arctic Tundra. The seed germinated and flowered after an est imated record of 10,000 years of dormancy. A recent record of 2000 years old viable seed is of the date plam, Phoenix dactylifera discovered during the archeological excavation at King Herod's palace near the Dead Sea.
18. The endosperm is formed after the fertilization in Angiosperms . It is triploid while in Gymnosperm , endosperm is formed before the fertilization and haploid in nature .
19 . Normally , Dicotyledon embryonic development is Cruifer-type which is also known as onagrad-types.
20 . The ovules of some parasitic plants donot have integument e. g. Sandal [Santalum] and Viscum .
21 . The polysiphonous condition is found in families of Malvaceae and cucurbitaceae .
22 . Those plants which live immortal through the seeds are known as therophytes .
23 . The synergids are absent in the embryosac of Plumbago .
24. The pollin ation takes place at two ce lled structure of pollen grains in most of Angiosperms.
25. Monospor icem bryo sac extensively found in Angiosperm. It is discovered by Strasburger in Polygon umdivarictum . It develops only from one functional me gaspore.
2 6 . The seed of Cuscuta and Santalum lacks of cotyledon
2 7 . The middle layer is absent in Wolffia 28. Costum and Nico dia plants have mult ilayere d tape tum.
2 9 . The dormancy is absent in the seed of most of Crop plants and M an g roove plants like Rhi zop hora .
3 0 . The fru it fo rmed from the ovary wit hout fert iliza tion is c alle d "P arthen oca rpic fruits " .
31. Lotus, Nymphaea and Alis ma like aquatic plants are entomophilous.
3 2 . All the microspores of anther of Calotropis are joined together to form a special, structure called Pollinium .
These staminal structures are called tr a nslat or ap par at us .
3 3 . In some pla nt s like Typh a, Dros era, Cr yptoste gi a al l fou r microspore s joi ned toget he r a nd t he y are cal led Compound pollen.
3 4 . For the formation of mature pollen grain, One meiotic and one mitotic divisions required. For the formata ti o n of mature male gametophyte, one meiotic and two mitotic divisions required.
35. In orc his macu lata and Ru mex , T-s hape d tetrad of megas pore is f ormed (In mos t angi osperms g enera lly linear letrads are formed. Isobilateral, tetrahedral and decussate type tetrad of megaspores are absent). 36. Balanoph o ra and Oe nothera plants one mi cropy lar meg aspore of tetrad bec omes func tional res t of three which are present towards the chalaza degenerate.
3 7 . When entry of male gametes into the embryosac are not participate in fusion is called Semigamy .
3 8 . When two pollen tube enter into an ovule and release their contents. It is poss ible to that the one male gamete of one pollen tube fertilize with egg cell and one male gamete of another pollen tube participate in triple fusion (with secondary nucleus). It is called heterofertilization . Because of this xenia effect found in Zea mays .
3 9 . The white fluid is found in green coconut is nuclear endosperm which is known as "liquid sync ytiumonlyonlyonly
40. In Cerea ls [Monocots] outer layer of endosperm is thick walled with dense cytoplasm, which is mainly filled with ale urone g rai n s [h ighly protei n rich ] a nd hence cal le d Aleuro ne L aye r . T h is l ayer secre te so me hy droly t ic enzymes like pr ot e ases a nd amylases .
I. DEFINITION OF DORMANCY OF SEED
The presence of dormancy in a seed is the mos t important characteristic feature of plants. Because of this character, seeds remain v iable for ma ny years. The seeds are dispersed ver y far place s through water, air or insects. Most of the seeds are unable to germinate just after dispersal. They germinate after sometime. The time between the maturation and germination of seed is known as "Dormancy period". The state of inhibited germination as a result of internal causes is usually called 'dormancy' . This seed dormancy is of considerable advantage to the plant which helps to pass on adverse environmental conditions. The em bryo remains in ac tive in this perio d an d all the growt h pro cess es su spend ed te mpora rily . The re are three mai n ba sic reas ons of dormancy of seed :-
II. REASONS OF SEED DORMANCY
1 . Impermeability of s eed c oat The seed coats of many species of Leguminosae and Convolulaceae families are c ompletely impermeable to mois ture [water] and oxygen at the time of their maturity. S eed co ats, in these are t hick and ha rd . Their cell wall is covered by a layer of lignin which is water proof coating. Such seeds take more time for germination.
Under natural c onditions seed dormancy is gradually over c ome by the action of microbes [bac teria] in soil, in the alimentary canal of fruit eater birds and due to the presence of low or high temperature.
SEED - DORMANCY
2 . Dormant Emb ryo In many spec ies, although, the embryo completely not matured when the seed is ripe, even than it fails to germinates w hen ever the envi ronmental conditions are favoura ble or even seed coat is removed. This is known as "Emb ryo " d or manc y" . Normally embryo dormancy occurs in many fr u its y ielding p lant s of forest. This condition is achieved due to physiological action of seed. Such type of seed must complete their enzym atic and chemi cal reac tion before the g ermination of seed. In t he lack of these reactio ns, seeds are una ble t o ge rmina te. s uch s eed a re k ept i n low t emp erature an d in de sir able m ois ture . Due to this s eed dormancy can be broken. The low temperature is the main reason for germination of seeds of Apple, Peaches, Pea rs , Map ple an d P ine etc.
3. Germination Inh ibitors Germination of some seeds is sometimes checked or preven ted by the presence of some chemical compounds are called "Germi nat ion Inh ibitors " such as Feru li c ac id pre sent in Tomato juic e , Caumarin, Abscisic Ac id, Dormi n and Para ascorb ic acid etc . T his c ondit ion is gene rally foun d in x er o phytic pla n ts . The se germination inhibitors washed away with water is known as "Leaching".
III. METHODS OF BREAKING OF SEED DORMANCY
(i) Scarification :- The hard seed coat is broken in this method so that water and oxygen enter into the seed coat. The dormancy of seeds can be broken by different artificial methods -
(i) By making minute hole/pores with help of pointed sharp apparatus on the seed coat.
(ii) The seeds are rubbed on hard object so that seed coat become thin.
(iii) The partial degeneration of seed coat is carried by the action of sulphuric acid.
(ii) Stratific ation :- The seed kept at low tempr a ture and in the presence of oxygen and water for some time, so that embryo can completes its maturation.
(iii) Lig ht Requirement :- The seeds which are affected by the lig ht are known as " Photoblastic seeds ".
The seeds which germinate in the presence of light are called "positive photoblastic seeds", such as Lettuce , Capsella , Lepidium and Tobacco etc. T he seeds i n which germinat ion take place in the absence of light are called "negative photoblastic seeds" , such as Ni g ella and Silene etc. Some seeds are not affec ted by light are called "nonphotoblastic seeds".
The dormanc y of photoblastic seeds can be broken by the treatment of red light . The phytochrome red , absorbs red light and convert into ph ytoc hrome far red which increase the germination of seed.
Seed + R [red light] → Germination Seed + R + FR → No germination Seed + R + FR + R → Germination Seed + R + FR + R + FR → No germination
(iv) Alte rnat ive Te mp erat ure :- Seeds are treated with high and low temperature alternatively at high pressure to increase the germination of seed. e.g.The seed of Malilotus (Sweet clover) and Alfa - alfa are treated at 2000 atm os pheric pres sure and 18 0 C temperature.
(v) Viabi lity :- This is called existanc e of l ife in a s eed . The viability of seed can be tested out by 2, 3 , 5, triph enyl te tr a zo li um chl or id e . The embryonal ax is of living seed becomes pinkin colourin the solution of T.T.C.
(vi) Seed dormancy can also be broken by the treatment of G.A.
IV. SPECIAL POINTS
(1) Hard and imperme ab le se ed coat is found in most of legume plantshich is the ma in reason of seed dormancy. A hard lignin water resistant layer [seed coat] is present on seed of some Leguminoseae plants.
(2) Growth inhibitor is pres ent in the endo sperm of Ir is , in seed coat of Cucurb ita and in the embryo of X ant hium .
(3) The seed dormancy in Eranthis he ema lis and Gi nk go is p re se nt d ue to imm at ure emb ryo .
(4) Some chemic alshave been used success fully to in terruptseed dormancy caused by resistan torhard impermeable seed coat. such as potassium nitrate , Ethylene , Chlrohy drineand Thiourea etc.
( 5 ) Atriplex seed contai ns high concentration of salt of the time of dispersal, whi ch do not allow ge rmination.
BUD DORMANCY
Before showing their respective growth, the bud of many plants undergoes a dormant phase. This is known as bud dormancy. This period starts in late summer and terminate in the sprin g sea so n.
CAUSES OF BUD DORMANCY -
According to Hemberg in woody plant, the bud dormancy is caused by Abscisic ac id. The level of endog enous A BA increases with the onset of dormant period and decreases when it is broken. In non w oody plant like Pota to , the bud dorma nc y is agai n d ue to the pre sen ce of a n i nh ibitor. B a n ne t Cl ark a nd Keffo rd (19 53) ident ified t h is subs ta nce as Inh ibito r-b . It p re sent i n t he peel of dorma nt Pota to tub er . T he i nh ibitor-b is re sponsible for checki ng t he sprout i ng of buds located i n t he 'eye s' of Potato tube r.
Accordi ng to Wareing. The bud dormancy, at least in woody plant, is caused by shor t day length, wh ile it is broken by long day.
BREAKING OF BUD DORMANCY
( i ) Chilling : The bud dormanc y of some plant can be broken, if they are given cold temperature treatment for specific duration.
( ii ) Alternating temperature treatment : The dormancy of buds of some plants can be broken, if they are subjec ted to low temperature (0°-10°C) for a brief d uration a nd then gi ven w a r m t rea t m en t .
( iii ) High temperature treatment : If dry P ota to tub e rs are stored at 35° C or moist at 20°C, the dormancy of tubers buds is broken.
( iv ) Chemic als : Some c hemicals l ike 2-c hlor o et hano l , Gib bere llin and T h io u rea are capable of breaking of dormanc y of buds . Of these 2-chloro ehtan ol is very effective in breaking the dormancy of P otato tu be r's buds . End ogen ous G ibberelli n play s a very signifi cant ro le in co ntrolli ng the d ormancy of P ota to t ube r s .
Chemicals : Some chemicals like 2-chloro ethanol, Gibberellin and Thiourea are capable of breaking of dormancy of buds. Of these 2-chloro ethanol is very effective in breaking the dormancy of Potato tuber's
buds. Endogenous Gibberellin plays a very significant role in controlling the dormancy of Potato tubers.
FAQs on Reproduction in Plant, Biology, Class 12
| 1. What is plant reproduction? |  |
| 2. What are the different modes of plant reproduction? | |
| 3. How do plants reproduce sexually? | |
| 4. What is asexual reproduction in plants? | |
| 5. Why is plant reproduction important? | |

|
4.5K Views |

|
4.62/5 Rating |

|
Dec 22, 2024 Last updated |

|
Explore Courses for Class 12 exam
|
|
Class 12
,Extra Questions
,Sample Paper
,ppt
,Viva Questions
,Summary
,Free
,practice quizzes
,Class 12
,MCQs
,Class 12
,past year papers
,Reproduction in Plant
,Biology
,Biology
,Reproduction in Plant
,Important questions
,shortcuts and tricks
,Reproduction in Plant
,video lectures
,Objective type Questions
,Biology
,Previous Year Questions with Solutions
,mock tests for examination
,Exam
,Semester Notes
,study material
;






Reproduction in Plant, Biology, Class 12 Free PDF Download
Importance of Reproduction in Plant, Biology, Class 12
Reproduction in Plant, Biology, Class 12 Notes
Reproduction in Plant, Biology, Class 12 Class 12 Questions
Study Reproduction in Plant, Biology, Class 12 on the App
|
© EduRev
|
Education Revolution
|



|




