NEET Exam > NEET Notes > Biology Class 11 > Important Diagrams: Photosynthesis in Higher Plants
Important Diagrams: Photosynthesis in Higher Plants | Biology Class 11 - NEET PDF Download

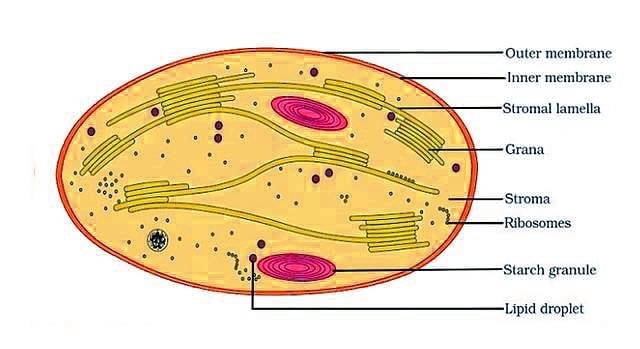
The Structure of a Chloroplast
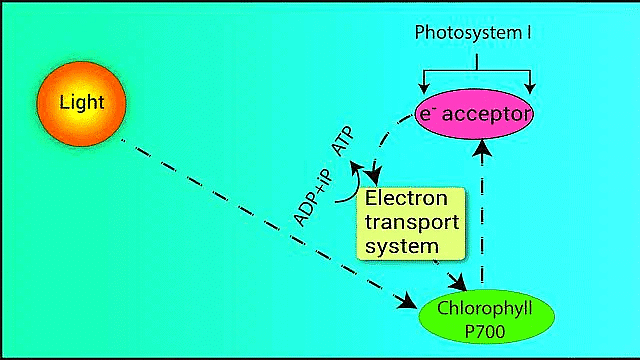
Cyclic Photophosphorylation
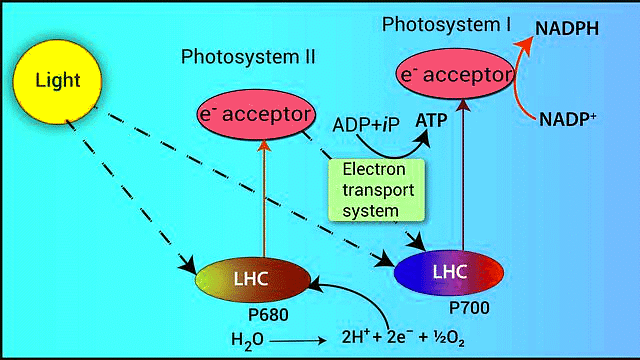
Non-cyclic Photophosphorylation
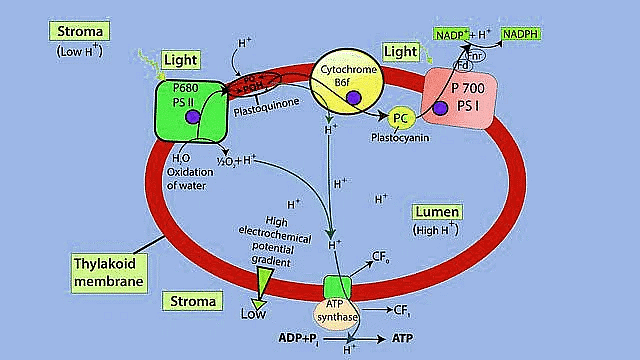
Electron Transport System
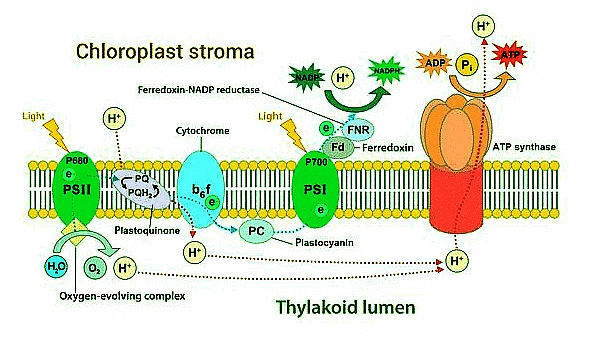
Chemiosmotic Hypothesis
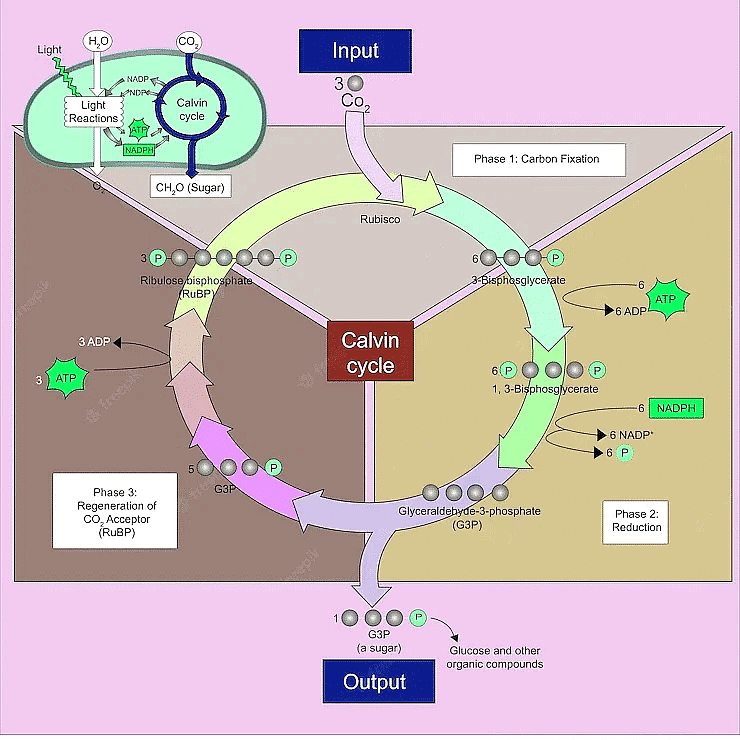
Calvin Cycle
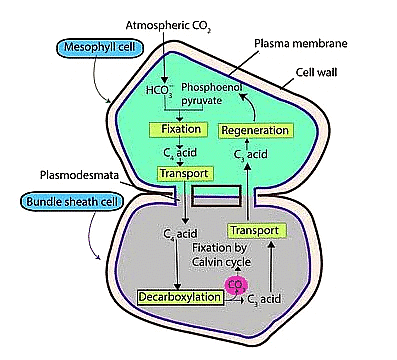
Hatch & Slack Cycle in C4 Plants
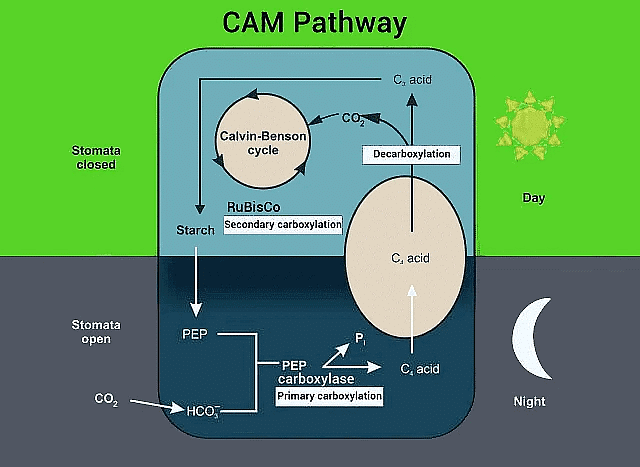
CAM Pathway
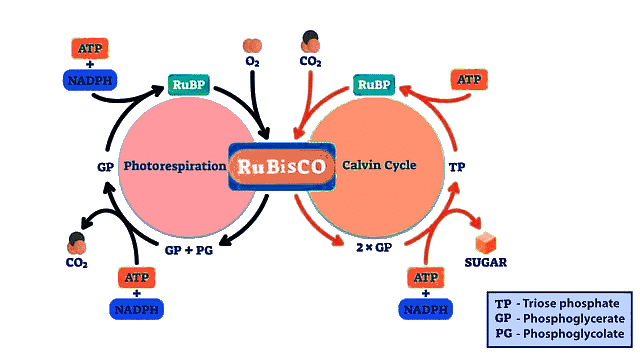
Photorespiration
Pigments involved in Photosynthesis
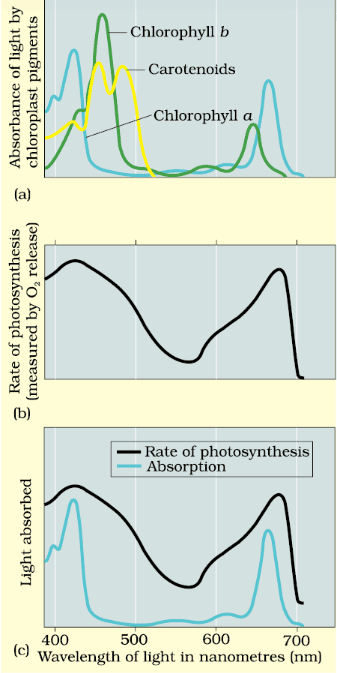 (a) Graph showing the absorption spectrum of chlorophyll a, b and the carotenoids (b) Graph showing action spectrum of photosynthesis (c) Graph showing action spectrum of photosynthesis superimposed on absorption spectrum of chlorophyll a
(a) Graph showing the absorption spectrum of chlorophyll a, b and the carotenoids (b) Graph showing action spectrum of photosynthesis (c) Graph showing action spectrum of photosynthesis superimposed on absorption spectrum of chlorophyll a
Light Reaction
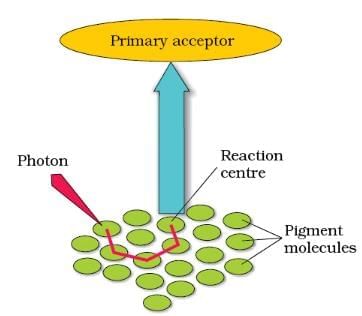 Light Harvesting Complex
Light Harvesting Complex
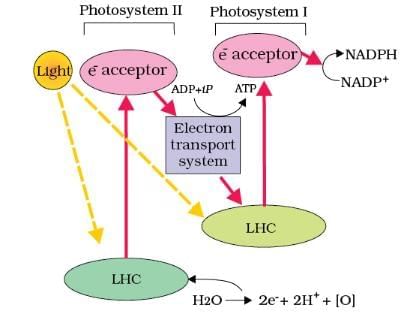 Z Scheme of Light Reaction
Z Scheme of Light Reaction
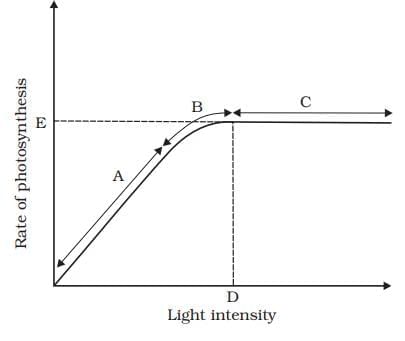
Light Effect on Photosynthesis
The document Important Diagrams: Photosynthesis in Higher Plants | Biology Class 11 - NEET is a part of the NEET Course Biology Class 11.
All you need of NEET at this link: NEET
|
150 videos|399 docs|136 tests
|
FAQs on Important Diagrams: Photosynthesis in Higher Plants - Biology Class 11 - NEET
| 1. What is the role of Joseph Priestley's experiment in understanding photosynthesis? |  |
Ans. Joseph Priestley's experiment in 1771 demonstrated that plants can produce oxygen. He used a candle in a sealed jar, which went out due to a lack of oxygen. When he introduced a green plant into the jar, the candle could be reignited, indicating that the plant released oxygen during the process of photosynthesis.
| 2. Where specifically in the plant does photosynthesis take place? | |
Ans. Photosynthesis primarily occurs in the chloroplasts of plant cells. These organelles contain chlorophyll, the green pigment that captures light energy and facilitates the conversion of carbon dioxide and water into glucose and oxygen.
| 3. How many types of pigments are involved in photosynthesis, and what are their functions? | |
Ans. There are two main types of pigments involved in photosynthesis: chlorophylls and accessory pigments. Chlorophyll a and b are the primary pigments that absorb light energy, while accessory pigments like carotenoids and xanthophylls help capture additional light wavelengths and protect the plant from excess light.
| 4. What is the light reaction in photosynthesis? | |
Ans. The light reaction, also known as the photochemical phase, occurs in the thylakoid membranes of the chloroplasts. It involves the absorption of sunlight by chlorophyll, leading to the splitting of water molecules (photolysis), the release of oxygen, and the generation of energy-rich molecules ATP and NADPH, which are used in the subsequent dark reactions.
| 5. What is the Calvin cycle, and how does it contribute to photosynthesis? | |
Ans. The Calvin cycle, also known as the dark reaction or light-independent reaction, takes place in the stroma of the chloroplasts. It utilizes ATP and NADPH produced in the light reactions to convert carbon dioxide into glucose through a series of enzymatic reactions, ultimately contributing to the plant's energy storage and growth.
About this Document

1.2K Views

4.85/5
Rating

Oct 10, 2025
Last updated
Document Description: Important Diagrams: Photosynthesis in Higher Plants for NEET 2025 is part of Biology Class 11 preparation.
The notes and questions for Important Diagrams: Photosynthesis in Higher Plants have been prepared according to the NEET exam syllabus. Information about Important Diagrams: Photosynthesis in Higher Plants covers topics
like The Structure of a Chloroplast, Cyclic Photophosphorylation, Non-cyclic Photophosphorylation, Electron Transport System, Chemiosmotic Hypothesis, Calvin Cycle, Hatch & Slack Cycle in C4 Plants, CAM Pathway, Photorespiration, Pigments involved in Photosynthesis, Light Reaction, Light Effect on Photosynthesis and Important Diagrams: Photosynthesis in Higher Plants Example, for NEET 2025 Exam. Find important definitions, questions, notes, meanings, examples, exercises and tests below for Important Diagrams: Photosynthesis in Higher Plants.
Introduction of Important Diagrams: Photosynthesis in Higher Plants in English is available as part of our Biology Class 11
for NEET & Important Diagrams: Photosynthesis in Higher Plants in Hindi for Biology Class 11 course.
Download more important topics related with notes, lectures and mock test series for NEET
Exam by signing up for free. NEET: Important Diagrams: Photosynthesis in Higher Plants | Biology Class 11 - NEET
Description
Full syllabus notes, lecture & questions for Important Diagrams: Photosynthesis in Higher Plants | Biology Class 11 - NEET - NEET | Plus excerises question with solution to help you revise complete syllabus for Biology Class 11 | Best notes, free PDF download
Information about Important Diagrams: Photosynthesis in Higher Plants
In this doc you can find the meaning of Important Diagrams: Photosynthesis in Higher Plants defined & explained in the simplest way possible. Besides explaining types of
Important Diagrams: Photosynthesis in Higher Plants theory, EduRev gives you an ample number of questions to practice Important Diagrams: Photosynthesis in Higher Plants tests, examples and also practice NEET
tests

Related Searches
ppt
,practice quizzes
,video lectures
,Previous Year Questions with Solutions
,shortcuts and tricks
,Viva Questions
,Free
,past year papers
,Important Diagrams: Photosynthesis in Higher Plants | Biology Class 11 - NEET
,Semester Notes
,Summary
,Objective type Questions
,Sample Paper
,Important Diagrams: Photosynthesis in Higher Plants | Biology Class 11 - NEET
,Important Diagrams: Photosynthesis in Higher Plants | Biology Class 11 - NEET
,mock tests for examination
,Extra Questions
,MCQs
,Exam
,study material
,Important questions
;


Additional Information about Important Diagrams: Photosynthesis in Higher Plants for NEET Preparation
Important Diagrams: Photosynthesis in Higher Plants Free PDF Download
The Important Diagrams: Photosynthesis in Higher Plants is an invaluable resource that delves deep into the core of the NEET exam.
These study notes are curated by experts and cover all the essential topics and concepts, making your preparation more efficient and effective.
With the help of these notes, you can grasp complex subjects quickly, revise important points easily,
and reinforce your understanding of key concepts. The study notes are presented in a concise and easy-to-understand manner,
allowing you to optimize your learning process. Whether you're looking for best-recommended books, sample papers, study material,
or toppers' notes, this PDF has got you covered. Download the Important Diagrams: Photosynthesis in Higher Plants now and kickstart your journey towards success in the NEET exam.
Importance of Important Diagrams: Photosynthesis in Higher Plants
The importance of Important Diagrams: Photosynthesis in Higher Plants cannot be overstated, especially for NEET aspirants.
This document holds the key to success in the NEET exam.
It offers a detailed understanding of the concept, providing invaluable insights into the topic.
By knowing the concepts well in advance, students can plan their preparation effectively.
Utilize this indispensable guide for a well-rounded preparation and achieve your desired results.
Important Diagrams: Photosynthesis in Higher Plants Notes
Important Diagrams: Photosynthesis in Higher Plants Notes offer in-depth insights into the specific topic to help you master it with ease.
This comprehensive document covers all aspects related to Important Diagrams: Photosynthesis in Higher Plants.
It includes detailed information about the exam syllabus, recommended books, and study materials for a well-rounded preparation.
Practice papers and question papers enable you to assess your progress effectively.
Additionally, the paper analysis provides valuable tips for tackling the exam strategically.
Access to Toppers' notes gives you an edge in understanding complex concepts.
Whether you're a beginner or aiming for advanced proficiency, Important Diagrams: Photosynthesis in Higher Plants Notes on EduRev are your ultimate resource for success.
Important Diagrams: Photosynthesis in Higher Plants NEET Questions
The "Important Diagrams: Photosynthesis in Higher Plants NEET Questions" guide is a valuable resource for all aspiring students preparing for the
NEET exam. It focuses on providing a wide range of practice questions to help students gauge
their understanding of the exam topics. These questions cover the entire syllabus, ensuring comprehensive preparation.
The guide includes previous years' question papers for students to familiarize themselves with the exam's format and difficulty level.
Additionally, it offers subject-specific question banks, allowing students to focus on weak areas and improve their performance.
Study Important Diagrams: Photosynthesis in Higher Plants on the App
Students of NEET can study Important Diagrams: Photosynthesis in Higher Plants alongwith tests & analysis from the EduRev app,
which will help them while preparing for their exam. Apart from the Important Diagrams: Photosynthesis in Higher Plants,
students can also utilize the EduRev App for other study materials such as previous year question papers, syllabus, important questions, etc.
The EduRev App will make your learning easier as you can access it from anywhere you want.
The content of Important Diagrams: Photosynthesis in Higher Plants is prepared as per the latest NEET syllabus.
|
© EduRev
|
Education Revolution
|



|






Signup to see your scores
go up
within 7 days!
within 7 days!
Takes less than 10 seconds to signup