Molecular Basis of Inheritance Chapter Notes | Biology Class 12 - NEET PDF Download

The DNA (Deoxyribonucleic acid)
DNA is the genetic material in most of the organisms except some viruses, which have an RNA genome, e.g. TMV (Tobacco mosaic virus). RNA mostly acts as a messenger, and an adapter and has a catalytic function. The number of base pairs (bp) or nucleotides determines the length of the DNA.
Human DNA (haploid) – 3.3 x 109 bp
Bacteriophage 𝜙 x 174 – 5386 nucleotides
Bacteriophage 𝝀 – 48502 bp
E. coli – 4.6 x 106 bp
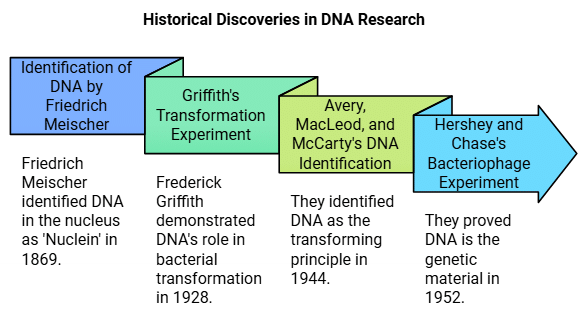
- Friedrich Meischer in 1869 identified DNA present in the nucleus as an acidic substance and named it as ‘Nuclein’
- Frederick Griffith in 1928 demonstrated in the infected mice that some “transforming principle” gets transferred from heat-killed S-strain of Streptococcus pneumoniae to R-strain, enabling it to make the smooth polysaccharide coat, hence it becomes virulent
- Oswald Avery, Colin MacLeod and Maclyn McCarty determined the biochemical nature of “transforming principle” and discovered that only DNA is responsible for the transformation
- Hershey and Chase in 1952 proved that DNA is the genetic material. They infected the bacteria E. coli with bacteriophages grown in radioactive phosphorus (32P) which labels DNA of the bacteriophage, which gets transferred to the bacteria cells and radioactive sulfur (35S), which labels the protein coat of the bacteriophage, hence radioactivity is not detected in the bacterial cell
Structure of a Polynucleotide Chain
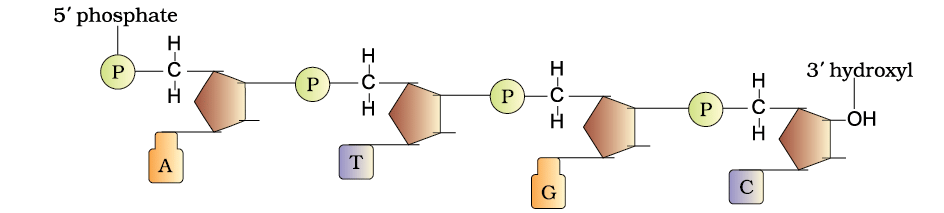
Each nucleotide is composed of three elements:
(i) Nitrogenous base:
- Purines- Adenine (A) and Guanine (G) present in DNA as well as RNA
- Pyrimidines- Cytosine (C) and Thymine (T) in DNA and Cytosine and Uracil in RNA. Thymine is also known as 5-methyl uracil and it is accounted for more stability of DNA molecule
(ii) Sugar: Pentose sugar- Ribose in RNA (ribonucleic acid), deoxyribose in DNA
(iii) Phosphate group
- Nucleoside: a nitrogenous base linked to the hydroxyl group of 1’ C of the pentose sugar by N-glycosidic bond
- Nucleotide: phosphate group is attached to the hydroxyl group present at 5’ C of the nucleoside by a phospho-ester bond
- 3’-5’ phosphodiester bond links two nucleotides together to form dinucleotide and the chain continues to grow to form a polynucleotide
Double Helix Model for Structure of DNA
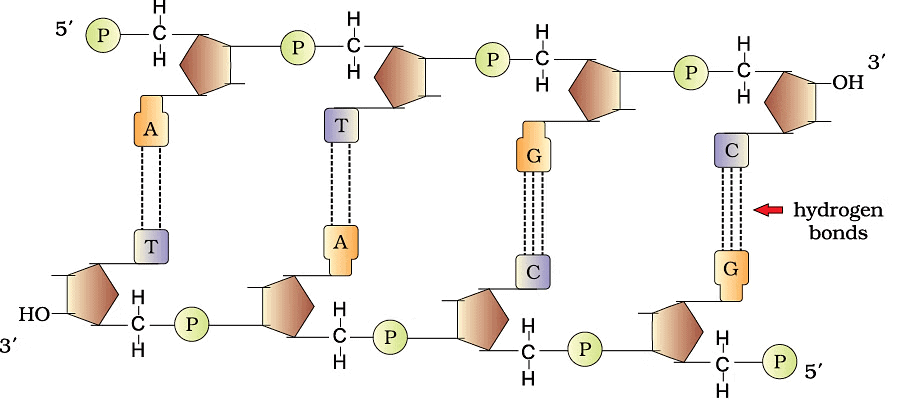
Watson and Crick in 1953 proposed the double helix structure of DNA.
Ervin Chargaff observed that the ratio of Adenine and Thymine to Guanine and Cytosine is one and remains constant
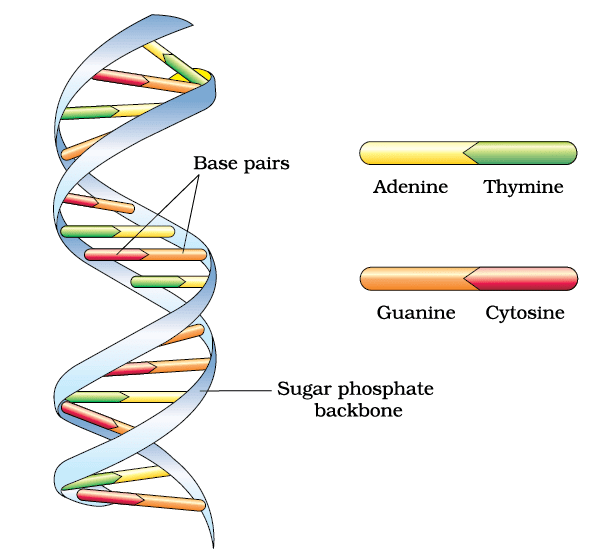
- Two polynucleotide chains make DNA, where the backbone is of sugar-phosphate and bases are present towards inside
- The two chains have the opposite polarity, i.e. one with 5’→3’ and the other has 3’→5’ polarity
- Base pair (bp) is formed by hydrogen bonding between nitrogenous bases of both the polypeptide chains
- Always a purine base of one nucleotide chain is linked to a pyrimidine base of another nucleotide chain or vice versa to make a base pair
- Adenine pairs with Thymine (or Uracil in RNA) by two hydrogen bonds (A=T)
- Guanine pairs with Cytosine by three hydrogen bonds (G)
- The two polypeptide chains are coiled in a right-handed direction
- The distance between two base pairs is 0.34 nm, there are 10 bp in each turn and the pitch of the helix id 3.4 nm
- Stability of the helical structure is conferred by the presence of base pairs one above the other
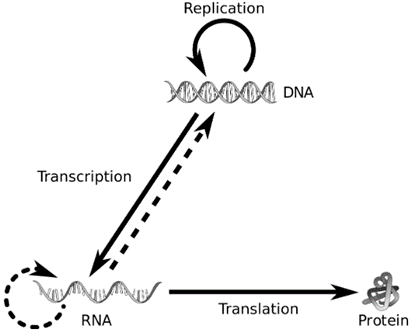
The proposition of a double helix structure for DNA and its simplicity in explaining the genetic implication became revolutionary. Very soon, Francis Crick proposed the Central dogma in molecular biology, which states that the genetic information flows from DNA->RNA->Protein.
Packaging of DNA Helix
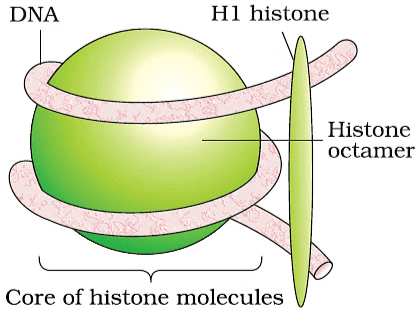
In prokaryotes, DNA is organised as a large loop in the nucleoid region. Negatively charged DNA is held together by positively charged proteins. In eukaryotes there occurs a complex organisation of DNA in chromosomes. DNA is wound around the core of histone octamer (a unit with 8 histone molecules) to form a nucleosome.
Histones are positively charged proteins as they are rich in basic amino acids; lysine and arginine
- There are 5 types of histone proteins- H1, H2A, H2B, H3 and H4
- Histone octamer has 2 molecules of 4 histone proteins and it plays an important role in gene regulation
- A nucleosome is a repeating unit in chromatin, it prevents DNA from getting tangled
- Nucleosome contains around 200 bp of DNA
- Non-histone chromosomal proteins (NHC) help in further packaging of chromatin
- Euchromatin: these are transcriptionally active areas, where chromatin is loosely packed and they take up light stain
- Heterochromatin: these are transcriptionally inactive areas, where chromatin is densely packed and they take up dark stain.
The Search for Genetic Material
Friedrich Miescher discovered nuclein, laying the groundwork for understanding DNA, even before Gregor Mendel's principles of inheritance. Although Miescher's findings and Mendel's principles were contemporaneous, it took time to establish DNA as the genetic material.
Early 20th Century Research: By 1926, research into the mechanism of genetic inheritance had advanced to the molecular level, focusing on chromosomes within the cell nucleus. Scientists like Gregor Mendel, Walter Sutton, and Thomas Hunt Morgan had narrowed down the search for genetic material to the chromosomes, but the specific molecule responsible for genetic inheritance remained unidentified.
Frederick Griffith's Experiment (1928)
- Griffith conducted experiments with two strains of Streptococcus pneumoniae to understand how bacteria could change their physical form.
- He observed that some bacteria produced smooth colonies (S strain) due to a polysaccharide coat, while others produced rough colonies (R strain) without this coat.

- Griffith found that when he injected heat-killed S strain bacteria into mice, they did not die. However, when he mixed heat-killed S strain with live R strain bacteria, the mice died.

- This led Griffith to conclude that some "transforming principle" from the heat-killed S strain was transferred to the R strain, enabling it to synthesize a smooth coat and become virulent.
- Griffith's experiments suggested the transfer of genetic material but did not define its biochemical nature.
Biochemical Characterization of the Transforming Principle
- Oswald Avery, Colin MacLeod, and Maclyn McCarty conducted further research to identify the biochemical nature of the transforming principle in Griffith's experiment.
- They believed the genetic material could be a protein and purified various biochemicals from heat-killed S cells to see which could transform live R cells into S cells.
- Through their experiments, they discovered that only DNA from S bacteria could transform R bacteria into S bacteria.
- They found that protein-digesting enzymes and RNA-digesting enzymes did not affect transformation, indicating that the transforming substance was neither a protein nor RNA.
- Digestion with DNase inhibited transformation, suggesting that DNA was responsible for the transformation.
- Avery and his colleagues concluded that DNA is the hereditary material, although not all biologists accepted this idea at the time.
- DNA carries genetic instructions for the growth, development, and reproduction of living organisms. It consists of two strands forming a double helix, made up of nucleotides containing a sugar, phosphate group, and nitrogenous bases. DNase is an enzyme that breaks down DNA by cutting the bonds between nucleotides, often used in labs to remove DNA or study its degradation. In Griffith's experiments, DNase's inhibition of transformation showed that DNA was the genetic material responsible for the change.
The Genetic Material is DNA
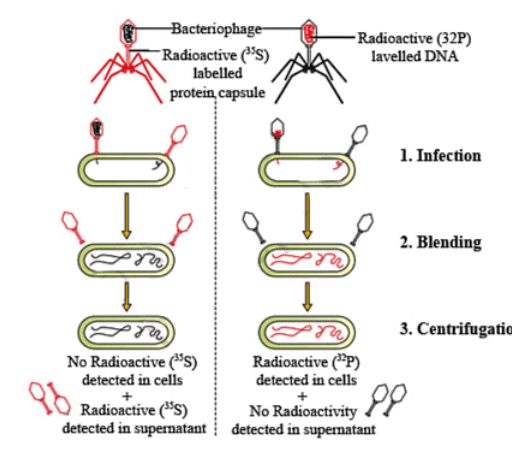
Hershey and Chase's Experiment (1952)
Hershey and Chase conducted experiments with bacteriophages (viruses that infect bacteria) to determine whether DNA or protein was the genetic material. Bacteriophages attach to bacterial cells and inject their genetic material, which the bacteria then use to produce more virus particles.
Radioactive Labeling: To differentiate between DNA and protein, Hershey and Chase labeled the genetic material of viruses using radioactive isotopes. They used radioactive phosphorus (³²P) to label DNA, since DNA contains phosphorus but protein does not. They used radioactive sulfur (³⁵S) to label protein, since DNA does not contain sulfur.
Infection of Bacteria: The radioactive phages were allowed to infect E. coli bacteria. After a period of infection, the viral coats were removed from the bacteria by agitating the mixture in a blender. The mixture was then spun in a centrifuge to separate the empty viral coats from the infected bacteria.
Results
(i)Bacteria infected with phages containing radioactive DNA (³²P) were found to be radioactive, indicating that DNA had entered the bacterial cells.
(ii) Bacteria infected with phages containing radioactive protein (³⁵S) were not radioactive, indicating that protein did not enter the bacterial cells.
Conclusion
(i)The results of the experiment provided strong evidence that DNA, not protein, is the genetic material that is passed from viruses to bacteria.
(ii) This experiment was crucial in establishing DNA as the genetic material in living organisms.
Properties of Genetic Material (DNA versus RNA)
The debate on whether DNA or proteins carried genetic information was resolved by the Hershey-Chase experiment, proving DNA as the genetic material. In some viruses (e.g., Tobacco Mosaic virus, QB bacteriophage), RNA serves as the genetic material instead of DNA.
- Genetic material must: Replicate itself , Be chemically and structurally stable & Allow gradual mutations for evolution.
- Express traits ('Mendelian Characters')
- Both DNA and RNA can replicate due to base pairing, while proteins cannot.
- DNA is stable and remains unchanged across life stages, as demonstrated by Griffith’s transforming principle.
- DNA's two complementary strands can reassemble after separation, contributing to its stability.
- RNA's 2'-OH group makes it chemically reactive and less stable, unlike DNA which is more structurally stable.
- DNA's use of thymine (instead of RNA's uracil) further enhances its stability.
- RNA is more prone to mutations and evolves faster, especially in RNA viruses.
- RNA can directly code for proteins, while DNA relies on RNA for protein synthesis.
- DNA is preferred for long-term genetic storage due to its stability; RNA is better for transmitting genetic information.
RNA World
RNA was likely the first genetic material. There is evidence to suggest that essential life processes, such as metabolism, translation, and splicing, evolved around RNA. RNA acted as both genetic material and a catalyst in early life forms. However, because RNA is reactive and unstable as a catalyst, DNA eventually evolved from RNA with chemical modifications that enhanced its stability. DNA, being double-stranded with complementary strands, is also better at resisting changes and has developed repair mechanisms.
Replication
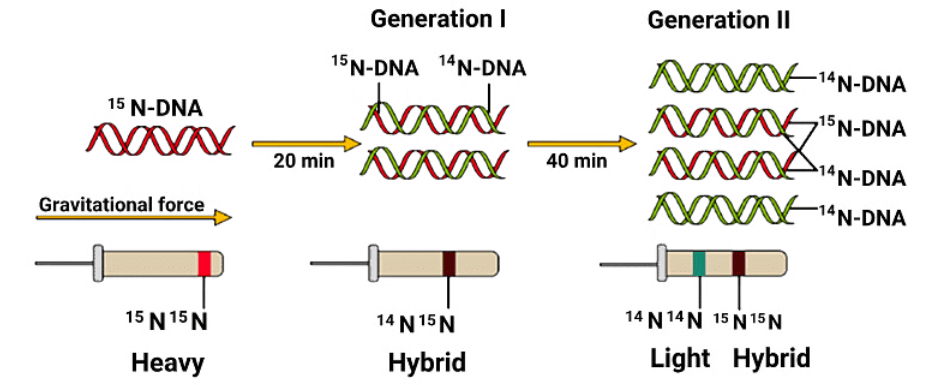
Watson and Crick suggested that the replication of DNA is semiconservative. Meselson and Stahl in 1958 experimentally proved that the DNA replicates semi conservatively. Taylor et al in another experiment on fava beans (Vicia faba) using radioactive thymidine proved that the replication on DNA is semiconservative
An experiment by Meselson and Stahl that demonstrated the semi-conservative replication of DNA,
Experiment Setup: Matthew Meselson and Franklin Stahl grew E. coli in a medium containing the heavy isotope of nitrogen, 15N, which was incorporated into the newly synthesized DNA.
Centrifugation Method: They used centrifugation in a caesium chloride (CsCl) density gradient to differentiate between the heavy DNA (with 15N) and the normal DNA (with 14N), based on their densities.
Transfer and Sampling: The E. coli were then transferred to a medium containing the normal 14N isotope, and samples were taken at various intervals as the bacteria multiplied, continuing to extract DNA which remained double-stranded.
Observations: After one generation (20 minutes), the extracted DNA showed a hybrid or intermediate density. After another generation (40 minutes), the DNA consisted of equal amounts of hybrid and light DNA.
Further Growth: By extending the experiment to 80 minutes, it was expected to find a mix of light and hybrid DNA densities, with an increasing proportion of light DNA as the number of generations increased.
Comparison and Proof: Similar experiments using radioactive thymidine on Vicia faba (fava beans) by Taylor and colleagues in 1958 corroborated the findings that DNA replicates semi-conservatively in chromosomes as well.
The Machinery and the Enzymes
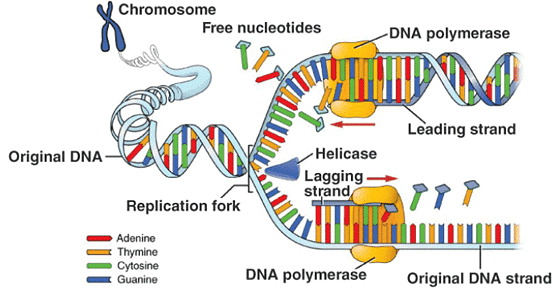
Enzyme DNA polymerase catalyses DNA replication. It can polymerise only in 5’→3’ direction
- Replication is initiated at the origin of replication
- Deoxyribonucleoside triphosphate provides energy for the polymerisation reaction and also acts as a substrate
- A small part of DNA opens up making a replication fork, where replication occurs
- Replication is continuous in a strand with 5’→3’ direction, called leading strand, where the template strand has 3’→5’ polarity, called leading strand template
- Replication is discontinuous in the other strand, where the template strand has 5’→3’ polarity, called lagging strand template
- The discontinuous fragments, called Okazaki fragments are joined together by the enzyme DNA ligase
- In eukaryotic cells, the replication takes place during s-phase of the cell cycle
- If cell division doesn’t occur after the replication, it results in polyploidy of chromosomes
Transcription
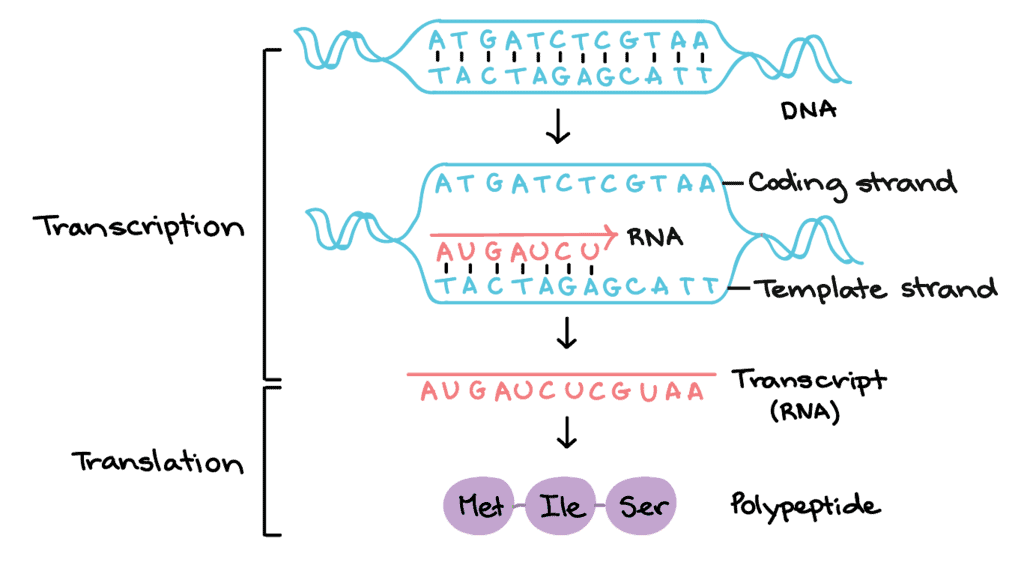
In the process of transcription, the genetic information present in the DNA gets copied to RNA. Only one segment of DNA gets copied to RNA. In RNA Uracil is present instead of Thymine in DNA.
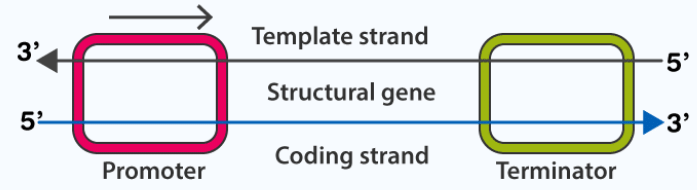
- Transcription of DNA includes three regions; a promoter, the structural gene and a terminator
- RNA polymerase catalyses the transcription and direction of transcription is same as that of replication by DNA polymerase, i.e. 5’→3’ direction
- Template Strand: it has 3’→5’ polarity, which acts as a template for RNA formation, also called antisense strand
- Coding strand: it has 5’→3’ polarity, which has the sequence similar to the newly formed RNA, except that Thymine is replaced by Uracil in RNA, also called the sense strand
- Promoter: it is situated at the 5’ side of the structural gene or upstream (with respect to coding strand). RNA polymerase binds here to start the transcription
- Structural gene: the region between promoter and terminator. Cistron is a segment of DNA that codes for a polypeptide. The structural gene is monocistronic in eukaryotic cells and polycistronic in prokaryotic cells
- Terminator: it is situated at the 3’ side of the coding strand and marks the end of the transcription process
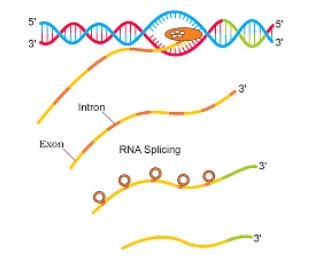
- The monocistronic structural gene (in eukaryotes) has interrupted coding sequences, the gene in eukaryotes is called split genes
- Exons: coding sequences, which appear in mature and processed RNA
- Introns: intervening sequences. It is not present in the mature and processed RNA
Transcription in a Prokaryotic cell
In bacteria, a single DNA-dependent RNA polymerase catalyses the transcription of all the three RNAs present; mRNA, tRNA, rRNA
There are three steps of transcription:
- Initiation: RNA polymerase binds to initiation factor, sigma (𝞂) at the promoter site to start the process of transcription
- Elongation: RNA polymerase is only capable of elongation
- Termination: when the RNA polymerase reaches the terminator region, it binds with the terminator factor, rho (𝜌) to terminate the process and the nascent RNA falls off
Transcription process in Bacteria
- In bacteria, mRNA doesn’t require further processing and as the nucleus and cytosol are not separate, translation is coupled with transcription and is initiated before the full transcription of mRNA occurs
Transcription in a Eukaryotic cell
There are three RNA polymerases to catalyse transcription of different RNAs in a eukaryotic cell:
- RNA polymerase I- transcription of rRNA (28S, 18S, 5.8S)
- RNA polymerase II- transcription of hnRNA (heterogeneous nuclear RNA), which is a precursor of mRNA
- RNA polymerase III- transcription of tRNA, 5S rRNA and snRNA (small nuclear RNAThe primary transcript is non-functional and has exons intervened by introns
Splicing- the process of removing introns and joining together exons in a defined sequence
Capping and tailing- additional processing of hnRNA. In capping a methyl guanosine triphosphate is added to the 5’ end of hnRNA. In tailing, 200-300 adenylate residues are added at 3’ end. mRNA is fully processed hnRNA, which gets transported out of the nucleus for translation.
Genetic Code
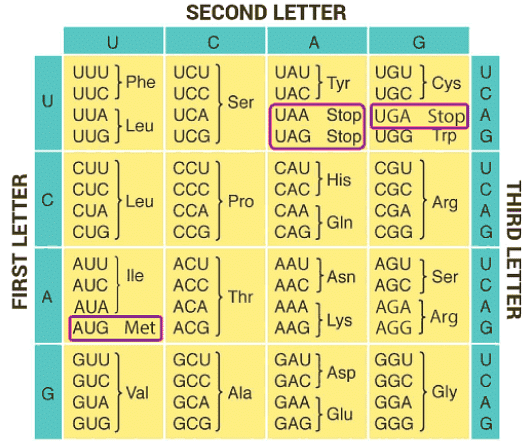
Genetic code is the sequence of bases in mRNA, that codes for a particular amino acid in the protein synthesis. Each code is made up of three nucleotides called a triplet. codons are nearly universal, except for some protozoans and mitochondrial codons. More than one triplet codon code for same amino acid, so the code is degenerate
- There are a total of 64 codons, of which 61 code for amino acids
- 3 codons do not code for any amino acids, they are called stop codons- UAA, UAG, UGA
- AUG is the start codon as well as codes for the amino acid methionine
Genetic Mutation
- Point Mutation: Change of single base pair results in the point mutation, e.g. Sickle cell anaemia is a result of a point mutation in the gene coding for the 𝛽-globin chain. As a result Glutamate in the normal protein gets converted to Valine in the sickle cell
- Frameshift Mutation: When there is loss or gain of one or two base pairs, it changes the reading frame at the point of insertion or deletion resulting in the frameshift mutation
Translation
The translation is the process of amino acid polymerisation. Amino acids are joined by peptide bonds
All three RNAs have a different role in the process of translation
- mRNA- provides the template. The sequence of amino acids in a polypeptide chain is determined by the sequence of bases present in mRNA
- tRNA- acts as an adapter, it brings amino acids and reads the genetic code
- rRNA- performs a structural and catalytic role
tRNA – The Adapter Molecule: Crick proposed the presence of an adapter molecule, which binds to a specific amino acid. It was known as soluble RNA or sRNA and later named as tRNA. Shape of the tRNA is like an inverted ‘L’. tRNA is specific for each amino acid, there is a specific initiator tRNA. There are no tRNAs for stop codons. It has an anticodon loop, which has complimentary code present on mRNA
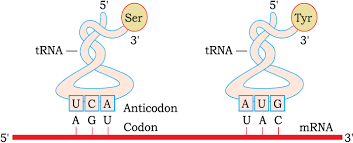
There is an amino acid acceptor arm, which binds the specific amino acid as per the codon
- The first step in the process of translation is aminoacylation of tRNA (charging of tRNA)
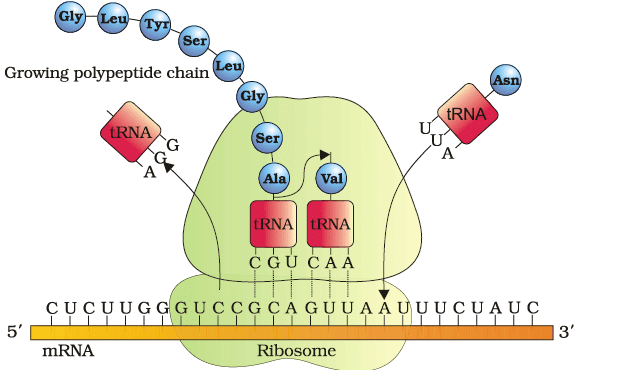
- Ribosomes are a protein manufacturing factory. mRNA to protein translation begins with the presence of mRNA in the small subunit of ribosomes
- The process of translation is always in 5’→3’ direction
- Peptide bond formation occurs between two amino acids present on tRNAs in close vicinity
- There are two sites in the large subunit of a ribosome which accommodates two tRNAs with amino acids close enough to form a peptide bond
- Ribosome also acts as a catalyst in the formation of a peptide bond
- A ribozyme is a 23s rRNA molecule present in bacteria, which acts as an enzyme catalysing peptide bond formation
- Start codon and stop codon flank the coding sequence for a polypeptide in mRNA
- Untranslated regions (UTRs)- UTRs are present before the start codon, i.e. at 5’ end and after the stop codon, i.e. towards 3’ end. They are not translated but they make the translation process efficient
- The Release factor binds to the stop codon at the end terminating the process. The polypeptide gets released from the ribosome
Regulation of Gene Expression
Expression of a gene to form polypeptide can be regulated at various levels in eukaryotes
- At the time of formation of a primary transcript, i.e. transcription
- At the time of processing or splicing
- At the time of transportation of mRNA from the nucleus to the cytosol
- At the time of protein synthesis, i.e. translation
Environmental, physiological and metabolic conditions regulate the gene expression. The development and differentiation of embryo is a result of coordinated regulation and expression of several sets of genes
In prokaryotes, control of gene expression is mainly at the initiation of transcription. The activity of RNA polymerase at the start site is regulated by regulatory proteins, which can be a repressor or activator
The accessibility of the promoter region is regulated by an operator sequence adjacent to it, that binds with the specific protein, mostly a repressor. In each operon, there is a specific operator and repressor protein
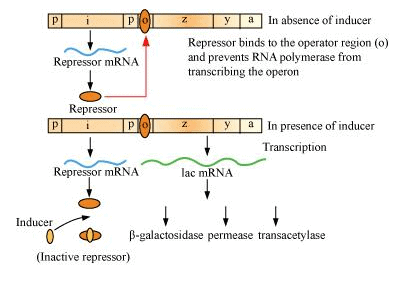
The Lac Operon
Jacob and Monad first showed the transcriptionally regulated system in the lac operon. An operon consists of many structural genes regulated by a common promoter and regulatory gene
The lac operon consists of
- Regulatory gene: gene i (inhibitor gene), that codes for the repressor of the lac operon
- Structural gene: three structural genes, z, y, a
- z gene, codes for 𝜷-galactosidase, it hydrolyses lactose to glucose and galactose
- y gene, codes for permease, responsible for increasing permeability of the cell to 𝛽-galactosides
- a gene, codes for transacetylase
The repressor is synthesised continuously from the gene i, which binds to the operator and prevents RNA polymerase from transcribing. Lactose is a substrate of 𝜷-galactosidase and also regulates the gene expression. It acts as an inducer. When lactose or allolactose is present, it binds with repressor and inactivates it, allowing RNA polymerase to access to the promoter region and initiating the transcription. Regulation by repressor in called negative regulation
Human Genome Project
Human genome project (HGP) was launched in 1990 to decipher the complete DNA sequence of the human genome. Genetic engineering techniques were used to isolate and clone the DNA segment for determining the DNA sequence. The project got completed in 2003, the sequence of chromosome 1 was completed in May 2006.
Some of the key findings of HGP are:
- The human genome has 3164.7 million bp
- Total ~30,000 genes are present with an average of 3000 bases per gene
- Dystrophin gene is the largest human gene having 2.4 million bases
- 99.9 % of nucleotides are the same in all people
- Only 2 % of genome codes for proteins
- Most genes are found on chromosome 1, i.e. 2968
- Least genes are found on the Y chromosome, i.e. 231
- There are around 1.4 million locations, where there is a single base difference in DNA, it is called single nucleotide polymorphism- SNPs (snips)
DNA Fingerprinting
- Alec Jeffreys initially developed the DNA fingerprinting techniques and named it as VNTR (Variable Number of Tandem Repeats)
- The difference in the DNA makeup accounts for the unique phenotype of each individual and that is the basis of DNA fingerprinting
- The DNA sequence of two individuals can be compared very quickly by the DNA fingerprinting technique
- DNA fingerprinting involves identifying the difference between two DNA molecule at the specific regions where the sequence is repeated many times called repetitive DNA
- These repetitive DNAs make a small peak during density gradient centrifugation and known as satellite DNA
- Depending on the number of repetitive units, base composition and length of the segment, satellite DNA is further classified into micro-satellites, mini-satellites, etc.
- These sequences do not code for protein but make a large portion of the human genome
- A high degree of polymorphism present in these sequences is the basis of DNA fingerprinting
- DNA fingerprinting is used for paternity test as this polymorphism is inherited to the child
- It has been widely used in forensic science
- DNA fingerprinting can be used in determining genetic diversity existing in a population
|
59 videos|290 docs|168 tests
|
FAQs on Molecular Basis of Inheritance Chapter Notes - Biology Class 12 - NEET
| 1. What is the primary function of DNA as genetic material? |  |
| 2. How does DNA differ from RNA in terms of structure and function? | |
| 3. What is the significance of the RNA world hypothesis in understanding the origin of life? | |
| 4. What are the main steps involved in the process of transcription? | |
| 5. How can genetic mutations affect protein synthesis and function? | |




Molecular Basis of Inheritance Chapter Notes | Biology Class 12 - NEET
,Objective type Questions
,Extra Questions
,Summary
,past year papers
,Important questions
,video lectures
,MCQs
,Semester Notes
,Sample Paper
,study material
,Viva Questions
,Free
,practice quizzes
,Exam
,Molecular Basis of Inheritance Chapter Notes | Biology Class 12 - NEET
,mock tests for examination
,ppt
,Molecular Basis of Inheritance Chapter Notes | Biology Class 12 - NEET
,shortcuts and tricks
,Previous Year Questions with Solutions
;


Chapter Notes: Molecular Basis of Inheritance Free PDF Download
Importance of Chapter Notes: Molecular Basis of Inheritance
Chapter Notes: Molecular Basis of Inheritance
Chapter Notes: Molecular Basis of Inheritance NEET Questions
Study Chapter Notes: Molecular Basis of Inheritance on the App
|
© EduRev
|
Education Revolution
|



|






within 7 days!