Pigments as photoreceptors | Botany Optional for UPSC PDF Download

Photoreceptors of Photosynthetic Pigments
Photosynthesis is a fascinating process that sustains life on Earth by converting sunlight into chemical energy. It involves a complex interplay of various pigments and photoreceptors. In this article, we will delve into the world of these photoreceptors, specifically focusing on seven main types of photoreceptors associated with photosynthetic pigments.
1. Phycobilins
Phycobilins are a group of pigments found in a range of photosynthetic organisms, including red algae, cyanobacteria, and green plants. These pigments serve as accessory light-harvesting molecules in photosynthesis. Four types of phycobilins have been identified, with three directly involved in photosynthesis and the fourth, known as phytochromobilin, regulating growth and development.
- Phycoerythin, phycocyanin, and allophycocyanin are the phycobilins involved in photosynthesis.
- Phycobilins consist of open-chain tetrapyrrole and an integral protein, forming a chromoprotein.
- These pigments are organized into structures called phycobilisomes.
- Phytochrome, a type of phycobilin found in higher plants, plays a crucial role in various photomorphogenic processes.
Phycobilins indirectly contribute to photosynthesis by absorbing light energy and transferring it to chlorophyll a, which utilizes it for photosynthesis. They are considered accessory pigments, along with chlorophylls b, c, and d.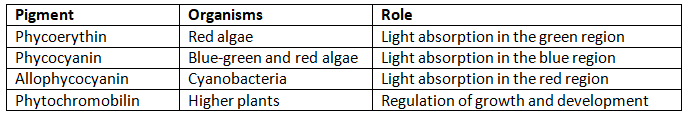
2. Cryptochrome
Cryptochromes are photoreceptors responsible for various responses to blue and UV-A radiation. These photoreceptors have long been a mystery, with carotenoids and flavins being suggested as potential candidates. Recent research in Arabidopsis has identified the HY4 gene as encoding the blue photoreceptor, CRY1-a cytoplasmic protein similar to photolyase. Cryptochromes play a crucial role in mediating responses to light, including the inhibition of hypocotyl elongation.
3. UV-B Receptors
Recent studies in plants like Sorghum vulgare and carrot cells have indicated the presence of UV-B receptors, although their exact nature remains elusive. These receptors seem to modulate responses to phytochrome in some systems.
4. Flavonoids
Flavonoids are responsible for the vibrant colors seen in flowers, such as scarlet, purple, pink, and blue. Anthocyanins, a type of flavonoid, give rise to these colors, while flavones are responsible for yellow flower pigmentation. Flavonoids are phenylpropane derivatives with a basic C6–C3–C6 composition. Three major groups of flavonoids are flavones, flavonols, and anthocyanidins. Anthocyanins are water-soluble pigments found in the vacuolar sap of plant cells and are known to protect against UV radiation.
5. Betacyanins
Betacyanins are complex glycosylated compounds responsible for the red pigments found in plants like beetroot and Bougainvillea bracts. Unlike some pigments, betacyanins do not respond reversibly to changes in pH, and they contain nitrogen within their molecules.
6. Chloroplasts
- Electron microscopy has provided insights into the structure of eukaryotic chloroplasts, which consist of an outer double-membrane layer without perforations, encasing a protein-rich interior called the stroma. The inner membrane extends within the chloroplast to create a system of lamellae that form stacks called grana. A mature chloroplast typically contains 40 to 60 grana. These membranes are paired, with thinner sections referred to as stroma lamellae. The pigments essential for photosynthesis are primarily located in the lamellar system within the chloroplast.
- The flattened membrane sacs within chloroplasts are called thylakoids, while the coin-shaped components within a granum are called grana thylakoids. Stroma thylakoids connect adjacent grana thylakoids, and their interior spaces are continuous with the thylakoid spaces within individual grana thylakoids. A granum can contain as many as one hundred stacked thylakoids. Additionally, chloroplasts contain darkly staining round granules called plastoglobuli, ribosomes, and aggregated DNA fibrils. Plastoglobuli are composed of substances like vitamin K and plastoquinones and are believed to play a role in membrane biosynthesis.
- Chloroplasts share some structural features with mitochondria, such as multiple membrane layers, the presence of DNA and ribosomes, and lipoprotein membranes with ADP phosphorylating systems. However, they differ in their function, number, and shape within a given cell, as well as their overall organization.
- In blue-green algae, thylakoids are distributed throughout the cytoplasm with regular spacing and are not enclosed within distinct organelles. They are situated peripherally and are rarely attached to membranes. In purple bacteria, thylakoids develop as deeply infolded regions of the plasma membrane and are continuous with the cell's outer membrane.
- The stroma of chloroplasts contains enzymes involved in the fixation of CO2 and its conversion into carbohydrates, fats, and proteins. It also houses some DNA and ribosomes. Chloroplasts can replicate independently of the cell nucleus, and their ribosomes can synthesize chloroplast proteins.
- Chloroplasts, also known as chromatophores, contain pigments that convert light energy into chemical energy during photosynthesis. There are three types of pigments in photosynthetic cells: chlorophylls, carotenoids, and phycobilins. Phycobilins are specific to red and blue-green algae and are found in phycobilisomes, distinct from chloroplasts.
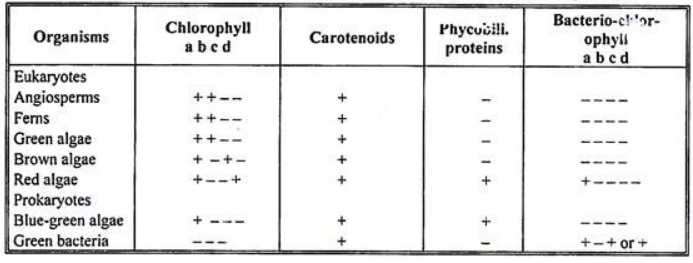
- Chlorophylls are the most crucial pigments for photosynthesis, with several types: chlorophylls a, b, c, d, and e; bacteriochlorophylls a, b, c, and d; and chlorobium chlorophyll (bacteriovirdin). Chlorophylls a and b are present in all autotrophic organisms except pigment-containing bacteria. Chlorophyll b is absent in blue-green, brown, and red algae. Chlorophylls c, d, and e are found only in algae and in conjunction with chlorophyll a.
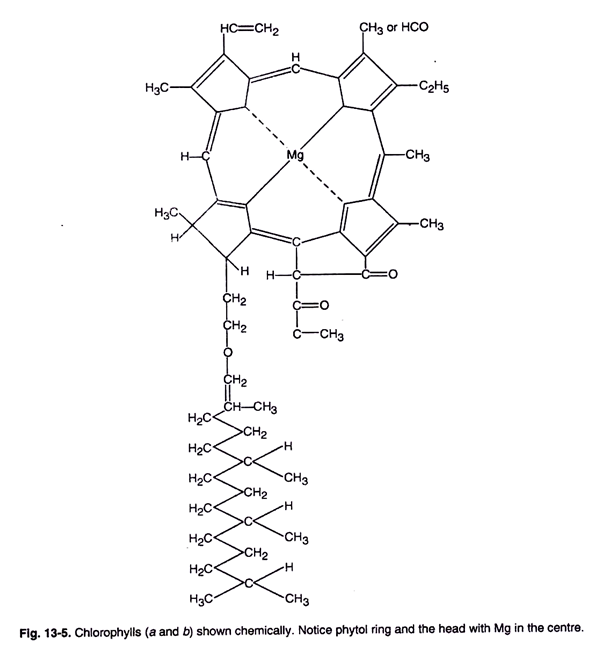
- The chlorophyll molecule has a cyclic tetrapyrrolic structure (porphyrin) with an isocyclic ring containing a magnesium atom at its center. Chlorophyll a appears blue-green, while chlorophyll b is yellow-green. They have different substitutions in the pyrrole ring of porphyrin.

- Chlorophyll a and b are soluble in different solvents, with absorption maxima in the blue-violet and red regions. They are highly fluorescent in solution but do not exhibit fluorescence in intact plants.
- The synthesis of chlorophyll a involves several steps, including the formation of aminolevulinic acid, porphobilinogen, uroporphyrinogen III, coproporphyrinogen-III, protoporphyrinogen IX, and finally, chlorophyllide a, which is converted to chlorophyll a. Various cofactors, enzymes, and essential minerals like iron, magnesium, and nitrogen are necessary for this process. Genetic factors also influence chlorophyll synthesis, and mutations can result in albino seedlings lacking chlorophyll.
Some of the suggested steps are as follow:
- Succinyl COA, an intermediate of Krebs cycle combines with glycine amino acid to form δ- amino β- Ketoadipic acid as unstable compound. This loses CO2 to yield aminolevulinic acid. The presence of cofactors pyridoxal phosphate and iron are essential. The enzyme δ- aminolevulinic acid synthetasecatalyses it. As mentioned earlier, iron deficiency causes chlorosis of young leaves. Light is shown to mediate the condensation of these two compounds.
- In the next step two molecules of δ-aminolevulinic acid condense, and the process is mediated by the enzyme δ-aminolevulinic acid dehydrase, to form porphobilinogen. In this reaction there is a fusion of two molecules.
- Then 4 molecules of porphobilinogen condense to form uroporphyrinogen III. Four ammonium ions are lost in this reaction and the process is mediated by the enzyme uroporphyrinogen-Isynthetase and uroporphyrinogen III cosynthetase.
- The four acetic acid substitutes of uroporphyrinogen-III yield coproporphyrinogen-III and the reaction is catalyzed by uroporphyrinogen decarboxylase.
- Under aerobic conditions, coproporphyrinogen-III, in the presence of coproporphyrinogen oxidative decarboxylase gives rise to protoporphyrinogen IX.
- Protoporphyrinogen IX undergoes oxidation and thus protoporphyrin IX is formed. It takes magnesium to form Mg protoporphyrin IX. Mg protoporphyrin methyl esterase catalyzes the addition of a methyl group of Mg protoporphyrin IX. It may be mentioned that the methyl group is donated by S-adenosyl methionine.
- In the next step, Mg protoporphyrin IX mono-methylester is converted to protochlorophyllide.
- A phytol group is added to protochlorophyllide to produce protochlorophyll. Once it was believed that protochlorophyll is the immediate precursor of chlorophyll a. However, recent evidences suggest that the immediate precursor of chlorophyll a is chlorophyllide a. When the etiolated seedlings are subjected to light, protochlorophyllide is reduced to form chlorophyllide a. The light is essentially required for this conversion.
- In the final step esterification of a phytol group to chlorophyllide a occurs and so chlorophyll a is produced. Enzyme chlorophyllase is involved in the process.
In gymnosperms, some ferns, and many algae, chlorophyll can be synthesized in die dark solely through enzymatic activity. On the other hand, it is believed that chlorophyll b is formed from chlorophyll a. Some of the minerals like manganse, potassium, zinc, copper, magnesium, iron, and nitrogen are essential for the synthesis of chlorophyll.
When absent or deficient they cause chlorosis. Chlorophyll formation is also dependent upon genetic factors as well. Absence of the gene(s) essential for its formation in the genetic constitution, produces seedlings from the germinating seeds which lack chlorophyll. These are known as “albinos”.
Steps in Chlorophyll Synthesis are summarized below:
7. Carotenoids
- Carotenoids are pigments responsible for the various colors seen in fruits and vegetables. They are divided into two chemical groups: carotenes and xanthophylls. Carotenes are orange-red, while xanthophylls are yellow. These pigments have 40 carbon atoms and play multiple roles in photosynthesis, including light absorption and protection against photooxidation.
- Carotenoids are synthesized through the mevalonic acid pathway and undergo several stages, including dehydrogenation and cyclization.
These are as follow:
- Tail-to-tail addition of two units of geranylgeranyl pyrophosphate to form 40C skeleton called phyton.
- In a series of steps, phyton is dehydrogenated to form lycopene.
- Lycopene undergoes cyclization.
- Xanthophylls are derived from carotenes by the oxidation.
Role of Carotenoids:
- The carotenoids perform functions in diverse processes including photosynthesis, phototropisms and protection against excessive light. Their role in photosynthesis appears to be secondary since tissues rich in carotenoids and lacking chlorophyll do not photosynthesize.
- It is also believed that light energy absorbed by them is transferred to chlorophyll and utilised in photosynthesis. When a plant is exposed to wavelengths of light absorbed exclusively by the carotenoids, a red fluorescence of chlorophyll a is observed.
- Action spectrum and absorption spectrum for the green alga (Ulva) shows that photosynthesis is appreciable in blue-green region at 480-500 nm wavelengths, indicating some transfer of energy from the carotenoids to chlorophyll.
- They also prevent destruction of chlorophyll from the degradative effects induced by excessive light and molecular oxygen (photooxidation).
- They may also act as photoreceptors for light causing phototropism.
In conclusion, photoreceptors and pigments are essential components of the photosynthetic machinery in plants and other photosynthetic organisms. Understanding these photoreceptors and pigments helps scientists unravel the intricate processes of photosynthesis and its regulation, shedding light on the fundamental mechanisms of life on Earth.
|
179 videos|143 docs
|
FAQs on Pigments as photoreceptors - Botany Optional for UPSC
| 1. What are photoreceptors of photosynthetic pigments? |  |
| 2. How do photosynthetic pigments act as photoreceptors? | |
| 3. What is the role of photosynthetic pigments in photosynthesis? | |
| 4. Which are the main photosynthetic pigments found in plants? | |
| 5. What happens to the absorbed light energy by photosynthetic pigments? | |


video lectures
,practice quizzes
,Extra Questions
,Viva Questions
,Previous Year Questions with Solutions
,Important questions
,mock tests for examination
,Exam
,Semester Notes
,Summary
,Free
,study material
,past year papers
,Pigments as photoreceptors | Botany Optional for UPSC
,Sample Paper
,Objective type Questions
,shortcuts and tricks
,Pigments as photoreceptors | Botany Optional for UPSC
,Pigments as photoreceptors | Botany Optional for UPSC
,ppt
,MCQs
;






Pigments as photoreceptors Free PDF Download
Importance of Pigments as photoreceptors
Pigments as photoreceptors Notes
Pigments as photoreceptors UPSC Questions
Study Pigments as photoreceptors on the App
|
© EduRev
|
Education Revolution
|



|





