Hearing System | Zoology Optional Notes for UPSC PDF Download
| Table of contents |
|
| Hearing System |
|
| Outer Ear |
|
| Ear canal |
|
| Middle Ear |
|
| Inner Ear |
|
| Organ of Corti |
|
| Bone Conduction System |
|
| Vestibulocochlear Nerve |
|

Hearing System
Hearing is the sense through which living organisms perceive and comprehend the surrounding acoustic environment, capturing sound vibrations in the atmosphere. It serves as the primary means by which various species respond to a specific range of physical vibrations. Human hearing enables the perception of speech and other acoustic events, along with the spatial detection and localization of sound sources in a 360° range. However, human hearing has limitations in terms of sensitivity to sound intensities and frequencies, and it allows for complete 360° spatial orientation only when there are no obstructing acoustic barriers nearby. Consequently, to optimize the potential of wearers' hearing capabilities in audio head- and helmet-mounted displays (HMDs), designers and acquisition personnel must possess a thorough understanding of human hearing's anatomy and physiology.
The process of hearing is termed audition, and the anatomical structure responsible for processing incoming acoustic stimuli is referred to as the hearing system or auditory system. The human hearing system comprises two ears on the left and right sides of the head, the vestibulocochlear nerve, and the central auditory nervous system (CANS), which includes auditory centers in the brain and connecting pathways in the brainstem. Each ear is further divided into three functional parts: the outer (external) ear, the middle ear, and the inner (internal) ear. The overall anatomical structure of the human ear, divided into three functional parts, is depicted in Figure 8-1, with a more schematic representation provided in Figure 8-10. The inner ear encompasses the vestibule, semicircular canals, and the cochlea, housing two sensory organs: the organ of balance and the organ of hearing. The organ of balance's components are found within the vestibule and semicircular canals, while the organ of hearing, the organ of Corti, is located in the cochlea. The proximity of the senses of hearing and balance leads to interactions between the two. Figure 8-1. Overall structure of the human ear
Figure 8-1. Overall structure of the human ear
The structures of the human ear are embedded in the temporal bone of the skull, with only the pinna (outer ear) protruding outside and being visible. The temporal bone is a dense bony structure on each side of the head, forming part of the cranium around the brain. The cranium comprises eight bones connected by sutures, while the skull includes 14 facial bones making up the facial skeleton. The overall structure of the skull and the locations of its constituent bones are depicted in Figure 8-2. Note that the sphenoid bone, though appearing as a plate, is actually a butterfly-shaped bone extending across the skull's width.
The ear canal's location in the temporal bone is marked in Figure 8-3, situated in the lower part of the bone. The middle and inner ears reside in the petrous portion of the temporal bone, offering protection to delicate ear structures. Besides housing ear components, the petrous portion features the internal auditory meatus, a canal through which the vestibulocochlear (8th cranial) and facial (7th cranial) nerves pass. The facial nerve provides sensory and motor innervation to the face.
The anterior portion of the temporal bone forms the temporomandibular joint (TMJ) by articulating with the mandible's condyle. The superior part is the squamous portion, attaching to the occipital and parietal bones. The posterior section is the mastoid portion, including the mastoid process—a bony ridge behind the pinna. The mastoid process serves as the typical attachment site for bone conduction hearing aids and vibrators during hearing testing. The condyle, located just in front of the visible part of the outer ear, is an effective location for placing bone vibrators in speech communication applications.
Outer Ear
The external ear comprises two primary components: the pinna, which is the external flange of the ear, and the ear canal. Both components are depicted in Figure 8-4. The ear canal concludes with the tympanic membrane, commonly known as the eardrum, acting as the boundary between the outer ear and the middle ear. The pinna extends from the side of the head at an angle ranging from 25° to 35° (with a mean value of 30°) to the occipital scalp, serving as a device to collect sound. The entrance to the ear canal is situated within the pinna, in front of the pinna flap. Directed towards the eardrum, the ear canal channels sound waves and safeguards the eardrum from external elements such as dust, small insects, and variations in temperature.
Pinna
The pinna, or auricle, is a roughly ovoid-shaped structure characterized by an irregular surface with numerous grooves and depressions. Humans possess two pinnae, one on each side of the head. Similar to most paired anatomical structures, these two pinnae exhibit variations in shape, groove patterns, and slight asymmetry in both vertical and horizontal planes. These differences, coupled with the simultaneous reception of acoustic signals by both ears, play a crucial role in sound localization and explain why localization mechanisms are not universally applicable across individuals. Consequently, the Head-Related Transfer Function of one person cannot be directly used by another person without adjustments. Figure 8-4. The outer ear and its main elements
Figure 8-4. The outer ear and its main elements
The internal framework of the pinna consists of a single piece of cartilage attached to surrounding tissues and covered with skin. Nerve fibers from the great auricular nerve and auriculotemporal nerve innervate the pinna, while the blood supply is provided by the posterior auricular and superficial temporal arteries. Ligaments and small muscles connect the pinna to the head, although humans have lost the ability to actively direct the pinna toward incoming sounds, a feature retained by some other species.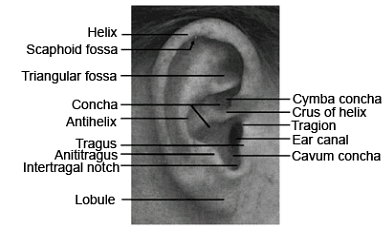 Figure 8-5. The pinna and its major structures
Figure 8-5. The pinna and its major structures
The average dimensions of the pinna are approximately 65 millimeters in length and 35 millimeters in width. In most adults, the width is approximately 55% of the length. The primary axis of the pinna points upwards with a posterior tilt of 5° to 30°, typically falling within the range of 15° to 20°. The highest point or superior aspect of the pinna is generally aligned with the brow.
The pinna exhibits prominent structures, including the helix (a curved rim), the antihelix (an internal ridge parallel to the helix), and the scaphoid fossa (a narrow curved depression separating the helix and antihelix). The concha, a major depression in the pinna, is divided into the cymba concha and cavum concha by the crus of the helix. The entrance to the ear canal is situated in the lower part of the pinna within the concha.
Notable features near the entrance to the ear canal include the tragus (a small cartilaginous flap covering the opening), the tragion (the notch above the tragus), and the antitragus (a cartilaginous flap below the tragus). The intertragal notch, located below the tragus and separating it from the antitragus, serves as a reference point for certain measurements. Additionally, the lobule (ear lobe), situated at the very bottom of the pinna, lacks cartilage and consists entirely of fat and skin.
Ear canal
The ear canal, also known as the auditory canal or external meatus, is a curved duct shaped like an "S" that serves as a pathway for acoustic waves to reach the tympanic membrane. Illustrated in Figure 8-4, the outer one-third of the ear canal is encased in cartilage, while the inner two-thirds enter the temporal bone, referred to as the cartilaginous and osseous parts, respectively. The cartilaginous segment is lined with a relatively thick layer of skin, continuous with that of the pinnae, and houses sebaceous glands, ceruminous (wax) glands, and hair follicles. Cerumen, or ear wax, is produced in the cartilaginous part, serving to moisturize the skin and trap dust and debris.
In contrast, the osseous part of the ear canal is covered with thinner skin, lacking hairs or secretion-producing glands. The skin in this region is innervated by branches of three cranial nerves: the auriculotemporal (mandibular) nerve, facial nerve, and vagus nerve. The outer layer of skin in the ear canal and tympanic membrane exhibits lateral migratory properties, facilitating self-cleaning by moving surface cells laterally from the tympanic membrane toward the ear canal opening. This migration occurs at an approximate rate of 100 microns per day.
The average length of an adult ear canal is around 25 mm, with a standard deviation of about 2 mm. Males typically have ear canals approximately 5% longer than females. The effective acoustic length is about 25% larger than its geometric length due to the "end effect" of the concha and its coupling to the ear canal. The canal is oval-shaped, with an average diameter of 7.0 to 8.0 mm. The cross-sectional area changes along its length, being approximately 0.45 cm2 at the opening and 0.4 cm2 in the middle. The last 8 to 10 mm of the ear canal is slightly tapered, reaching its narrowest point at the isthmus, located just past the second bend and about 4 mm from the tympanic membrane. The tympanic membrane terminates the ear canal at an oblique angle of 45° to 60°.
The ear canal's average volume ranges from approximately 1.0 to 1.4 cm3. Given that the proper selection and fitting of an audio Head-Mounted Display (HMD) depend on the dimensions of the human head, pinnae locations and dimensions, and the ear canal's geometry, mean values for these dimensions are provided in Tables 8-2 and 8-3 for reference. These dimensions are derived from large databases or anthropometric studies conducted with a significant number of participants.


Middle Ear
The middle ear is an air-filled cavity known as the tympanic cavity or tympanum. Its walls are constructed from the temporal bone and are lined with mucous membrane tissue. The total volume of the middle ear is approximately 2 cm3 (0.12 in3), as reported by Dallos (1973), Yost and Nielsen (1977), and Zemlin (1997). The lateral wall of the middle ear encompasses the tympanic membrane, as described earlier, while the medial wall is formed by a bony structure that separates the middle ear from the inner ear. This wall includes two membranous windows, the oval and round windows, establishing anatomical and physiological connections between the middle ear and the inner ear.
Maintaining a pressure just below atmospheric, the air in the middle ear cavity is regulated by the Eustachian tube (auditory tube), a narrow duct connecting the tympanic cavity to the upper part of the throat (nasopharynx). Within the middle ear cavity reside three small bones: the malleus (hammer), incus (anvil), and stapes (stirrup). Collectively known as the ossicles, these bones form an ossicular chain that links the tympanic membrane to the oval window. Suspended inside the cavity by middle ear ligaments and two middle ear muscles, the tensor tympani and the stapedius, this chain's structure is depicted in Figure 8-7.
 Figure 8-7. The middle ear and its main elements
Figure 8-7. The middle ear and its main elements
The tympanic cavity exhibits the shape of a narrow, irregular rectangular box, with a middle section narrower than its edges. Figure 8-8 provides a schematic representation of the middle ear cavity. The largest dimension of the tympanic cavity does not exceed 10 mm (0.4 in). Predominantly occupied by the ossicular chain and middle ear muscles, the cavity features a small empty space known as the epitympanum recess (attic) above the ossicles. The remaining vacant area is denoted as the tympanic cavity proper. The superior wall or ceiling of the tympanic cavity is formed by the tegmen tympani, a thin bone that separates the middle ear from the brain cavity.
A slender aperture called the aditus ad antrum, positioned at the top of the posterior wall, links the middle ear cavity to the mastoid antrum (tympanic antrum), a small chamber surrounded by mastoid air cells. The entrance to the Eustachian tube is situated in the anterior wall of the middle ear cavity. Forming the medial wall of the cavity are the oval and round windows of the inner ear, separated by a bony ridge known as the promontory. The inferior wall or floor of the cavity incorporates the jugular fossa, housing the jugular vein. The pulsation of blood in this vein can result in a ringing noise, known as tinnitus, in the ear.

Within the tympanic cavity, two of the smallest muscles in the human body, the tensor tympani muscle and the stapedius muscle, are situated. Among the two, the tensor tympani is relatively larger. The tendon of the stapedius muscle emerges from the pyramidal eminence of the posterior wall, connecting to the stapes. Another structure arising from the posterior wall is the chorda tympani nerve, a branch of the facial nerve. The chorda tympani traverses the middle ear from back to front, joining the lingual nerve and providing taste sensation to a section of the tongue. As for the tendon of the tensor tympani, it emerges from the anterior wall of the middle ear and attaches to the malleus bone. An anterior bone plate separates the middle ear from the internal carotid artery.
Tympanic membrane
The tympanic membrane, commonly known as the eardrum, is an oval, thin, semi-transparent membrane that marks the termination of the ear canal. Resembling a shallow cone, the membrane's tip, called the umbo, extends approximately 1.5 to 2 mm towards the middle ear and is attached to the manubrium ("handle") of the malleus bone. The apex angle of the tympanic membrane is around 120°. While most of the membrane is attached to the temporal bone surrounding the ear canal, there is a narrow area called the notch of Rivinus (incisura tympanica) where it is not attached. The dimensions along its major perpendicular axes are 9 to 10 mm and 8 to 9 mm, and the total surface area varies from 55 to 90 mm², with an average of 64 mm². Due to its conical shape, the effective area is approximately 55 mm². The tympanic membrane is thinnest at its center (about 70 μm) and thickest at its edge (30 μm to 120 μm). It comprises four layers: an outer epithelial layer continuous with the ear canal skin, two middle fiber layers responsible for membrane stiffness, and an inner mucus layer continuous with the middle ear lining. The outer layer is innervated by the auriculotemporal nerve. The membrane's average mass is approximately 14 mg. It has two main regions with different stiffness: a small lax triangular area called the pars flaccida and a larger, stiffer portion called the pars tensa. The pars flaccida compensates for small air pressure changes between the middle and outer ear, while the pars tensa is involved in transmitting acoustic energy from the outer ear to the middle ear. The modulus of elasticity (Young's modulus) for the tympanic membrane at its center is approximately 0.02-0.03 GPa.
Ossicular chain
The ossicular chain serves to link the tympanic membrane to the membrane covering the oval window. Its main function, along with the supporting muscles, is to convey sound energy from the tympanic membrane to the inner ear, while also providing protection for the inner ear against potentially damaging noise. A depiction of the ossicular chain along with the accompanying middle ear muscles is illustrated in Figure 8-9.
 Figure 8-9. The middle ear space and ossicle chain. Petrous part of the temporal bone.
Figure 8-9. The middle ear space and ossicle chain. Petrous part of the temporal bone.
The ossicles, which are the smallest bones in the human body, consist of the malleus, incus, and stapes. The malleus, the largest among them, measures approximately 8 to 9 mm (0.31 to 0.35 in) in length, while the stapes, the smallest human bone, is around 3 mm (0.12 in) long. The incus falls between them in size, ranging from 5 to 7 mm (0.20 to 0.28 in) in length. Their respective weights are approximately 25, 3, and 28 mg (0.00088, 0.00011, and 0.00099 oz). The malleus, positioned as the first bone in the ossicular chain, has its largest process, the manubrium (handle), firmly attached to the tympanic membrane. Both the manubrium and the head of the malleus move in unison. The incus connects to the malleus, with its body linked to the malleus head and its lenticular process connecting to the stapes, the final bone in the chain. The stapes, with its head, neck, arms, and footplate, forms a characteristic stirrup shape, and its footplate is tightly connected to the membrane covering the oval window.
The ossicles respond to acoustic pressure on the tympanic membrane and the actions of two middle ear muscles: the tensor tympani and stapedius muscles. These muscles contract to reduce the transmission of very intense sounds to the inner ear. The tensor tympani muscle, attached to the manubrium of the malleus, contracts to pull the handle of the malleus inward, regulating the tension on the tympanic membrane. On the other hand, the stapedius muscle, connected to the stapes, contracts to pull the stapes in an inferior and lateral direction, away from the oval window, limiting its range of motion.
Eustachian tube
The Eustachian tube is a thin tube, which connects the middle ear with the nose and throat (Figure 8-6). The tube is approximately 35 to 45 mm (1.4 to 1.8 in) long and it travels downward and inward from the middle ear to the nasopharynx (upper throat). At its upper end, the tube is narrow and surrounded by bone. Nearer the pharynx it widens and becomes cartilaginous.
The Eustachian tube is normally closed, opening only during swallowing and yawning. It is responsible for maintaining the air pressure within the middle ear at approximately ambient pressure. Similar pressure on both sides of tympanic membrane ensures that the tympanic membrane can vibrate maximally when struck by sound waves arriving from the ear canal.
Inner Ear
The inner ear is the final and the most complex part of the ear. It occupies a small bony cavity called the bony labyrinth (osseous labyrinth) that is located directly behind the medial wall of the middle ear. The inner ear consists of three main anatomical elements: the semicircular canals, the vestibule, and the cochlea. The structure and main elements of the inner ear are shown in Figure 8-10.
The bony labyrinth of the inner ear has a volume of approximately 2 cm3 and is lined by the membranous labyrinth that closely follows the shape of the bony labyrinth (Buckingham and Valvassordi, 2001). The blood supply to the membranous labyrinth is provided by various small blood vessels extending from the labyrinthine artery. The space between the bony labyrinth and the membranous labyrinth is filled with incompressible body fluid called perilymph.
The perilymph is high in sodium but low in potassium resembling in its chemical composition in the blood and the cerebrospinal fluid surrounding the brain. The space inside the membranous labyrinth is filled with another incompressible body fluid called endolymph. Endolymph is low in sodium but high in potassium and chemically resembles the intercellular fluid found inside cells in the body. The differences in the chemical composition of the perilymph and endolymph create an electric potential difference that like a battery, sustains the physiological activities of the sensory organs located in the inner ear.
 Figure 8-10. The inner ear and its main elements
Figure 8-10. The inner ear and its main elements
Functionally, the inner ear consists of two major elements: the cochlea and the vestibular system (comprising the utricle and saccule of the vestibule and the three semicircular canals). The cochlea contains the organ of hearing (organ of Corti) while the vestibular system contains five balance organs: two maculae (utricular macula and secular macula) and three cristae ampullares (one in each of the three semicircular canals). The cochlea and the semicircular canals are located at the two ends of the inner ear, while the utricle and saccule are parts of the centrally-located vestibule. The oval window is located on the wall of the vestibule and the round window is located at the base of the cochlea. The locations of the windows and the other major parts of the inner ear are shown schematically in Figure 8-11.
Two vary narrow channels; the vestibular aqueduct and the cochlear aqueduct (not shown in Figure 8-11); connect the inner ear with the cranial cavity surrounding the brain. At their narrowest points, the vestibular aqueduct and the cochlear aqueduct do not normally exceed 0.8 mm (0.03 in) and 0.15 mm (0.006 in) in diameter, respectively. Both aqueducts seem to have little effect on normal ossicular transmission of sound for frequencies above 20 Hertz (Hz) (see, for example, Gopen, Rosowski and Merchant [1997]) and their exact function is still unknown.
Cochlea
The cochlea is a coiled structure that resembles the snail and extends anteriolaterally from the vestibule. Its structural base is the bony spiral lamina, which makes 2½ to 2¾ turns around the bony core of the cochlea called the modiolus. The external diameter of the cochlea varies from approximately 9 mm (0.35 in) at its base to approximately 5 mm (0.20 in) at its apex (top) and its uncoiled length is 32 to 35 mm (1.25 to 1.38 in). The cochlea is divided along its length into three parallel channels: the scala vestibuli, scala media, and scala tympani.
The scala vestibuli and scala tympani are parts of the bony labyrinth whereas the scala media is a part of the membranous labyrinth. The inside of the cochlea is shown in Figure 8-12. The scala vestibuli and scala tympani are connected at the apex (top) of the cochlea through a small opening called the helicotrema. At the base of the cochlea, the scala vestibuli joins the vestibule. The scala vestibule is terminated at its base by the oval window, the fenestra ovalis, while scala media terminates at the round window, the fenestra rotunda. The membrane of the oval window has a surface area of 3.2 to 3.5 mm2 , is completely covered by the footplate of the stapes, and is sealed in the bony opening by the annular ligament. The round window has a surface area of approximately 2 mm2 and is located inferior and anterior to the oval window in the wall between the middle ear and inner ear and serves as a pressure valve between the two scalae. When an acoustic stimulus causes mechanical vibration of the stapes footplate this movement is translated to the membrane of the oval window. The membrane pushes back and forth on the perilymph of the scala vestibuli and through the helicotrema, the perilymph of the scala tympani. This motion results in alternating outward and inward movement of the membrane of the round window. The membrane bulges outward as the fluid moves from the scala vestibuli to the scala tympani and bulges inward as the fluid moves from the scala tympani to the scala vestibuli. The central most duct of the cochlea is the membranous scala media (cochlear duct). This duct separates the scala tympani and scala vestibuli. The border between the scala media and the scala vestibuli is Reissner’s membrane and the border between the scala media and the scala tympani is the basilar membrane. Reissner’s membrane (vestibular membrane) is attached to the osseous spiral lamina and projects obliquely from it to the outer wall of the cochlea forming a roof of the scala media. Together, the basilar and Reissner membranes would make the scala media into a closed tube if not for the ductus reunions, a tiny opening at its base that connects it to the saccule (Figure 8-11).The basilar membrane forms the floor of the scala media. The membrane is anchored to the spiral lamina on one end and to the spiral ligament on the other end. When uncoiled, the membrane has an approximate length of 32 to 35 mm (1.25 to 1.38 in), practically the same as the whole cochlea. Blood vessels and nerve fibers supporting the cochlea enter the cochlea through the modiolus and spiral lamina.
The central most duct of the cochlea is the membranous scala media (cochlear duct). This duct separates the scala tympani and scala vestibuli. The border between the scala media and the scala vestibuli is Reissner’s membrane and the border between the scala media and the scala tympani is the basilar membrane. Reissner’s membrane (vestibular membrane) is attached to the osseous spiral lamina and projects obliquely from it to the outer wall of the cochlea forming a roof of the scala media. Together, the basilar and Reissner membranes would make the scala media into a closed tube if not for the ductus reunions, a tiny opening at its base that connects it to the saccule (Figure 8-11).The basilar membrane forms the floor of the scala media. The membrane is anchored to the spiral lamina on one end and to the spiral ligament on the other end. When uncoiled, the membrane has an approximate length of 32 to 35 mm (1.25 to 1.38 in), practically the same as the whole cochlea. Blood vessels and nerve fibers supporting the cochlea enter the cochlea through the modiolus and spiral lamina.
The spiral lamina is narrower at the apex of the cochlea and wider at the base. Conversely, the basilar membrane is wider, thicker and more flaccid at the apex and narrower, thinner, and stiffer at the base. These factors affect the vibrational properties of the basilar membrane, which responds to low frequency vibrations at the apex and high frequency vibrations at the base of the cochlea.
The organ of hearing (organ of Corti) is located primarily on the basilar membrane, with a small segment projecting onto the spiral lamina. The organ of Corti is made up of sensory cells (hair cells) and supporting cells. A schematic cross section of the cochlea showing the content of the scala media and the structure of the organ of Corti are shown in Figure 8-13.

Organ of Corti
The organ of Corti converts the mechanical vibrations of the basilar membrane into neural impulses that then travel through the auditory nerve and brainstem to the brain. The organ is composed of sensory cells, called hair cells, and several types of supporting cells distributed along the length and width of the basilar membrane. The arrangement of the sensory cells and supporting cells on the basilar membrane is shown in Figure 8-13B. The fibers of the auditory nerves travel from the organ of Corti through a system of small perforations in the spiral lamina collectively called habenula perforata that start at the tympanic edge of the spiral lamina and continue further through it. From habenula perforata, nerve fibers travel through a channel in the center of the modiolus (Rosenthal’s canal), exit the base of the cochlea, and join vestibular nerve fibers to form the vestibulocochlear nerve.
There are two types of hair cells in the organ of Corti: the inner hair cells (IHCs) and the outer hair cells (OHCs). They are shown in Figure 8-14. Each hair cell has a number of small hair-like projections called stereocilia (cilia) extending from the top of the cell. The group of stereocilia at the top of a hair cell is called a stereocilia bundle. The stereocilia bundle of each hair cell is organized in several rows forming either a “W” or “V” pattern for OHCs and shallow “U” pattern for IHCs. Stereocilia in each row have graduated heights (like stair steps) and their tips are connected together by thin fibers called tip links. Each type of hair cell in the ear is connected to the nervous system by both afferent (ascending) and efferent (descending) nerve endings, but the number and function of these types of connections varies between IHCs and OHCs.
There are altogether approximately 3500 IHCs and approximately 12,000 OHCs distributed along each basilar membrane (thus, each ear contains between 15,000 and 16,000 hair cells). The IHCs are shaped like a flask and form a single row of cells supported by the spiral lamina. The OHCs have a cylindrical shape with a diameter of approximately 9 μm and are organized into three rows located farther away from the spiral lamina. The groups of IHCs and OHCs are separated by two rods (pillars) of Corti, which structurally support the organ of Corti. The inner rod rests on the spiral lamina while the outer rod is attached to the basilar membrane. The rods are attached at their tops and more widely separated at the base, forming a triangular shape called the tunnel of Corti. The tunnel is filled with the cortilymph fluid that has similar properties to the perilymph fluid found in the bony labyrinth. The tops of the hair cells and supporting cells of the organ of Corti are tightly connected together at their tips to form a continuous layer called the reticular lamina. The reticular lamina isolates all of the organ of Corti from the endolymph of the scala media except for stereocilia which project through the reticular lamina into the endolymph.
The OHCs are held in position by the outer rod of Corti on one side and by Deiters cells on the other side. Each Deiters cell holds an OHC at the bottom and through long projections called phalangeal processes from above. The middle part of an OHC is not firmly supported and is surrounded by a perilymph-filled space called the space of Nuel. Next to the Deiters cells, moving towards the outer end of the cochlea, there are several groups of supporting cells, called Hensen cells, Claudius cells, outer spiral sulcus cells, and Boettcher cells. Several of these cells are shown in Figure 8-13B. Lateral to these support cells is the Stria vascularis, a highly vascular organ attached to the outer surface of the scala media. Stria vascularis recycles potassium and produces endolymph for the scala media, thus maintaining the endocochlear potential (battery) of the inner ear.
The IHCs are structurally supported by the inner rod of Corti on one side and by inner sulcus and pharyngeal cells on the other (Lim, 1986). The inner sulcus cells occupy the region extending from IHCs toward the spiral limbus. The spiral limbus projects from the spiral lamina towards the organ of Corti and provides the attachment point for the tectorial membrane. The tectorial membrane is a gelatinous membrane extending above the organ of Corti from the upper surface of the spiral limbus. The largest stereocilia of the OHCs make contact with the tectorial membrane, and this connection is part of the mechanism that leads to the neural responses of the organ of Corti. The basic characteristics of the OHCs and the IHCs are summarized in Table 8-4.
The vestibular system groups together make up the peripheral organs of balance. The bony structures include three semicircular canals and the vestibule. Within the bony structure, the three semicircular canals contain the three membranous semicircular ducts and the vestibule contains the utricle and saccule. In evolutionary terms, the organs of balance are much older than the organ of hearing, which actually evolved from them (Zemlin, 1997). The arrangement of the components of the vestibular system is shown in Figure 8-11. The three semicircular canals are hoop-shaped structures connected at both ends to the vestibule. One of the ends is almost twice as wide as the other and is called the ampulla. The canals are perpendicular to each other and are called the horizontal (lateral), the anterior (superior), and the posterior (inferior) semicircular canal. The horizontal canal is located not exactly horizontally but forms a 30° angle relative to the horizon. The spatial arrangement of the canals is shown in Figure 8-15. Each semicircular canal is sensitive to head motion in the plane of that canal. The canals also form bilateral differential pairs between the ears (e.g., right anterior with left posterior which have their hair cells aligned oppositely). Rotation in one plane will be excitatory to one canal and inhibitory to the other.
The three semicircular canals are hoop-shaped structures connected at both ends to the vestibule. One of the ends is almost twice as wide as the other and is called the ampulla. The canals are perpendicular to each other and are called the horizontal (lateral), the anterior (superior), and the posterior (inferior) semicircular canal. The horizontal canal is located not exactly horizontally but forms a 30° angle relative to the horizon. The spatial arrangement of the canals is shown in Figure 8-15. Each semicircular canal is sensitive to head motion in the plane of that canal. The canals also form bilateral differential pairs between the ears (e.g., right anterior with left posterior which have their hair cells aligned oppositely). Rotation in one plane will be excitatory to one canal and inhibitory to the other.
The semicircular canals are filled with perilymph and the semicircular ducts are filled with endolymph. The ampulla (bulge) of each canal contains the crista ampulliaris, a saddle-shaped, raised section of wall that is populated with the hair cells whose stereocilia respond to angular acceleration. The stereocilia of the semicircular canals are embedded in a gelatinous mass called a cupula that is similar in function to the tectorial membrane of the organ of Corti, that is, its motion bends the hair cells, which then creates a neural impulse.
While the sensory organs in the semicircular canals respond to angular accelaration of the head, two other organs of balance, the sense organs within the utricle and the saccule, respond to gravity and linear acceleration in horizontal (utricle) and vertical (saccule) directions. The sense organs within the utricle and saccule are the maculae. They occupy the concave spaces at the bottom of the utricle and the saccule and contain tiny pieces of calcium carbonate, called otoliths (ear stones) or otoconia (ear dust), which are embedded into a gelatinous membrane (otolithic membrane) into which the stereocilia of the maculae project. Since the otoliths are quite numerous in the otolithic membrane and they are heavier then the surrounding fluid, the membrane gets displaced towards the Earth during head tilting (due to gravity) and away from the source of motion during linear acceleration (due to inertia); thus, head motions are translated by stereocilia deflection into neural impulses.
There are two types of hair cells in the semicircular canals and the vestibule. Type I hair cells are flask-shaped cells while type II hair cells are cylinder-shaped cells. Type I and type II hair cells are very similar in their structure and innervation to the inner hair cells and the outer hair cells of the organ of Corti, respectively. Each hair cell in the semicircular canals has 50 to 100 small stereocilia and a single larger cilium called a kinocilium, which only exists in rudimentary form in the hair cells of the cochlea. The stereocilia are arranged by length, with the longest stereocilia located close to the kinocilium, and are all connected by tip links. Movement of the stereocilia hair bundle toward the kinocilium causes a depolarizing (excitatory) sensory response whereas movement away from the kinocilium causes a hyperpolarization (inhibitory) sensory response.
Bone Conduction System
The inner ear receives mechanical vibrations and converts them into neural impulses by the organ of Corti. As such it can be stimulated by sounds transmitted through the system of outer and middle ear or by skull vibrations. The first mode of stimulation is called air conduction, while the second mode is referred to as bone conduction.
The anatomical system responsible for transmitting skull vibrations to the organ of Corti is called the bond conduction system.
The bone conduction system can be stimulated by either sound waves impinging on the human head or by delivering a vibratory signal by means of a mechanical driver (vibrator) coupled to the head. In the former case, the resulting stimulation is approximately 1000 times (60 decibels [dB]) weaker than the simultaneous air conduction stimulation. This is due to the air-bone mismatch when sound tries to enter the bones of the skull (Chapter 9, Auditory Function). Vibration of the whole head may also cause distractive interference between vibrations delivered to various parts of the skull. Therefore, such stimulation only has practical meaning in the case of people wearing hearing protectors or with severe conductive hearing loss (Henry and Letowski, 2007). In the case of people wearing audio HMDs or heavily soundattenuating head gear, even though their air conduction pathways may be completely blocked, they may still hear very intense sounds, such as explosions, jet engine sounds, pile driver impact sounds, etc. due to the stimulation received through bone conduction.
When the vibrations are delivered directly to the head by a mechanical driver, the driver-bone is much smaller than the air-bone mismatch and the person may hear even very weak vibrations of the driver. Thus, this mode of stimulation can be effectively used for speech communication, human-robot interaction, and delivery of tactical signals during military operations.
In order to understand the potential applications and limitations of bone conduction hearing it is necessary to understand the basic elements of the human head and how they interact with one another. The typical weight of the human head is approximately 3.5 kg and the basic dimensions of the male and female heads are given in Table 8-3. The head is a complex structure made of bones, cartilage, several types of soft tissue, and fluids (e.g., cerebrospinal fluid), which differ in their mechanical properties. These different forms of matter transmit sound with different speeds and with various degrees of attenuation. Densities for select components of the head together with their associated speeds of sound transmission are listed in Table 8-5. According to Evans and Lebow (1951) and Sauren and Classens (1993) the average density of the bones of the skull is 1412 kg/m3 , the Young modulus is 6.5 x 109 N/m2 , and the Poisson ratio2 is 0.22. All these values, however, are highly dependent on the amount of water in the bones and other tissue and the boundary conditions. The more dry and less constrained the bone is, the higher the speed of sound through the bone. In addition, solid matter, like the bone, can simultaneously propagate longitudinal, transverse (traveling), and surface waves that have different speeds and can interact with one another.
The bones of the skull are listed in Table 8-1 and their arrangement is shown in Figure 8-2. The manner in which the skull and associated tissue respond to mechanical stimulation depends on the point of stimulation and the frequency of the signal. Two typical driving points described in the literature are the mastoid process and the forehead. The distance from the mastoid process to the cochlea is approximately 30 mm (1.2 in) and is the shortest distance between the cochlea and the head periphery (Tonndorf and Jahn, 1981). The main stimulation pathway from the mastoid process lies wholly within the respective temporal bone, which results in relatively low attenuation of the initial stimulus. In addition, the direction of this stimulation is the same as that of the air conduction pathway, which causes elements of the latter pathway to also become excited by bone stimulation.
The forehead is the relatively flat surface of the frontal bone, which is the largest bone of the skull. It is a fairly symmetrical and deeply extended bone that can easily transfer its vibration to many other bones of the skull. Another effective bone conduction pathway in the skull is from the condyle of the mandible (jaw bone), which is located on the side of the head just in front of the entrance to the ear canal. Stimulation of condyle activates bones and cartilage surrounding the ear canal and tympanic cavity creates a secondary pathway through the air conduction system even more effectively than stimulation at the mastoid process. The most common excitation points for bone conduction communication are shown in Figure 8-3. Depending on the frequency of stimulation, the skull has several modes of vibration that differ in the phase relationship between the vibrations at different locations on the skull. At low frequencies, e.g. 200 Hz, the skull driven at the forehead location moves as a whole in a back and forth pattern (Bekesy, 1932). This type of vibration is called inertial vibration and its corresponding mode of vibration is called the inertial mode. At higher frequencies, e.g., 800 Hz, the direction of vibration remains the same, but the front and the back of the skull vibrate 180° out-of-phase. Vibration where different parts of the skull vibrate out-of-phase is referred to as compressional vibration. Out-of-phase vibration of the front and back of the head is called the first compressional mode. More complex compressional modes are elicited at even higher frequencies. Vibration patterns for the inertial and first two compressional modes of vibration are illustrated in Figure 8-16. A more detailed discussion of how different points and frequencies of stimulation affect bone conduction hearing, Auditory Function.
Depending on the frequency of stimulation, the skull has several modes of vibration that differ in the phase relationship between the vibrations at different locations on the skull. At low frequencies, e.g. 200 Hz, the skull driven at the forehead location moves as a whole in a back and forth pattern (Bekesy, 1932). This type of vibration is called inertial vibration and its corresponding mode of vibration is called the inertial mode. At higher frequencies, e.g., 800 Hz, the direction of vibration remains the same, but the front and the back of the skull vibrate 180° out-of-phase. Vibration where different parts of the skull vibrate out-of-phase is referred to as compressional vibration. Out-of-phase vibration of the front and back of the head is called the first compressional mode. More complex compressional modes are elicited at even higher frequencies. Vibration patterns for the inertial and first two compressional modes of vibration are illustrated in Figure 8-16. A more detailed discussion of how different points and frequencies of stimulation affect bone conduction hearing, Auditory Function.
Vestibulocochlear Nerve
The vestibulocochlear nerve (the VIII cranial nerve) is the nerve connecting the organs of hearing and balance to the brain. It consists of two parts: the auditory nerve (cochlear nerve) and the vestibular nerve. Sensory signals that produce sensations of sound travel from the ear to the brainstem via the auditory nerve. The ascending neurons of the auditory nerve innervate the hair cells of the organ of Corti, exit the organ of Corti through the habenula perforata, and then form the spiral ganglion in the modiolus. The ascending neurons, called the afferent neurons, carry information from the sensory cells toward the brain. The descending neurons, called the efferent neurons, carry information from the brain to the sensory cells and other cells of the peripheral nervous system.
A neuron consists of a neuron cell and input (dendrites) and output (axon) projections extending from the neuron cell. These projections are called nerve fibers. Depending upon their location and function, neurons may be any length from 1 inch to 4 feet (2.5 cm to 1.2 m) long. Neurons transmit electrochemical signals to and from the brain via their nerve fibers. Upon receiving a signal, one neuron sends information to its adjacent neuron, through a junction called a synapse
The nerve fibers of the auditory nerve originate all along the cochlea, from its apex to its base, and project to the cell bodies of these nerves, which form the spiral ganglion. The fibers extending from the apex follow a straight course and form the core of the spiral ganglion, while the fibers from the base are twisted to form the outside surface of the ganglion. After leaving the cochlea, the auditory nerve joins the vestibular nerve, another bundle of fibers supporting the vestibular system, and they together form a bundle of approximately 30,000 afferent and efferent nerve fibers called the vestibulocochlear nerve. The vestibulocochlear nerve exits the inner ear through the internal auditory meatus, approximately 1 cm long channel in the temporal bone, which also houses the facial nerve, and enters the brainstem, where the auditory and vestibular parts of the vestibulocochlear nerve separate and take different pathways through the central nervous system. The structural representation of the vestibulocochlear nerve is shown in Figure 8-17.
The ascending pathways of the vestibulocochlear nerve that support the hair cells of the organ of Corti involve two types of afferent neurons: inner radial neurons (type I afferent neurons) and outer spiral neurons (type II afferent neurons). The inner radial neurons constitute approximately 95% of the ascending neurons in the cochlea and the outer spiral neurons make up the remaining 5% (Gelfand, 1998). The inner radial neurons, which are myelinated (insulated with a fatty substance) and larger of the two, innervate the IHCs. The innervation pattern is many-to-one and approximately 8 to 10 afferent fibers supply one IHC (Gelfand, 1998). The outer spiral neurons, which are unmyelinated and thinner, innervate the OHCs. The innervation pattern is one-to-many with one neuron making synapse connections with approximately 10 OHCs (Gelfand, 1998). Similar to the ascending pathways, the descending pathways of the vestibulocochlear nerve also involve two types of efferent neurons. They are the lateral olivocochlear neurons and the medial olivocochlear neurons, both of which descend from the superior olivary complex in the brainstem. The lateral olivocochlear neurons are myelinated and the larger, more numerous of the two, and they synapse with the projections of the afferent neurons connected to the IHCs. They constitute approximately 20% of the efferent neurons in the cochlea. The remaining 80% of efferent neurons in the cochlea are medial olivocochlear neurons (Gelfand, 1998). They are thin and unmyelinated and synapse to the OHCs. The distribution of the efferent fibers on the OHCs heavily favors the base of the cochlea. A schematic view of the innervation pattern for IHCs and OHCs is shown in Figure 8-18.
Similar to the ascending pathways, the descending pathways of the vestibulocochlear nerve also involve two types of efferent neurons. They are the lateral olivocochlear neurons and the medial olivocochlear neurons, both of which descend from the superior olivary complex in the brainstem. The lateral olivocochlear neurons are myelinated and the larger, more numerous of the two, and they synapse with the projections of the afferent neurons connected to the IHCs. They constitute approximately 20% of the efferent neurons in the cochlea. The remaining 80% of efferent neurons in the cochlea are medial olivocochlear neurons (Gelfand, 1998). They are thin and unmyelinated and synapse to the OHCs. The distribution of the efferent fibers on the OHCs heavily favors the base of the cochlea. A schematic view of the innervation pattern for IHCs and OHCs is shown in Figure 8-18.
Central Auditory Nervous System
The central auditory nervous system (CANS) is a system of neural structures and connections within the brain that processes neural impulses transmitted from the vestibulocochlear nerve and converts them into auditory sensations. It is a subsystem of the central nervous system (CNS), which includes the entire brain and the spinal cord. The CNS is a dynamic system composed of various types of nerve cells (neurons), which form an extraordinary network of neural connections reaching out from the brain to every part of the body. The human brain is estimated to contain 100 billion (1011) neurons3 and a quadrillion (1015) synapses4 (Kimball, 2005). The anatomical organization of the main structural elements of the human brain is shown in Figure 8-19.
The most inferior (lowest) portion of the brain is the brainstem. The brainstem is approximately 10 cm long and 2.5 cm wide at the central core (Seikal, King, and Drumright, 2000). It is the superior extension of the spinal cord and the place where the vestibulocochlear nerve enters the brain. The main anatomical elements of the brainstem are the medulla oblongata, pons, and midbrain. The medulla oblongata, pons, and cerebellum form the posterior (back) part of the brain called the hindbrain. The midbrain is the most superior (top) part of the brainstem and it is connected to and located just below the forebrain (cerebrum), the largest and the most advanced part of the brain. The main parts of the forebrain are the telencephalon (including the cerebral hemispheres, basal nuclei, and medullary center of nerve fibers) and the diencephalons (including the thalamus, and hypothalamus).
Neural pathways in the CANS consist of various nuclei (groups of cell bodies) and fiber tracts (bundles of nerve fibers) which carry information between and among the nuclei. Each nucleus serves as a relay station for dispatching neural information from one nucleus to the next. The neurons comprising a specific neural pathway travel through several nuclei in the brainstem before reaching the auditory cortex. The nuclei involved in the classical ascending auditory pathway are the cochlear nucleus, superior olivary complex, inferior colliculus, and medial geniculate body. Neural fibers carrying specific information may synapse with nuclei on the same side or decussate (cross from one side to the other) and synapse with nuclei on the other side of the brainstem. The pathway that connects the nuclei on the same side of the brainstem is called the ipsilateral pathway and the pathway that crosses from one side to the other is called the contralateral pathway. A general view of the ascending auditory pathway is shown in Figure 8-20. Note the size of the lateral lemniscus, which is the largest fiber tract in the CANS. The cochlear nucleus, which spans the pons and medulla, is the first processing center and relay station of the ascending auditory pathways. The two other major relay stations in the brainstem are the superior olivary complex in the pons and the inferior colliculus in the midbrain. From the cochlear nucleus, the nerve fibers project to the superior olivary complex (SOC) or directly to the lateral lemniscus. Approximately 75% of the ascending CANS fibers leaving the cochlear nucleus cross over to the contralateral side of the brain to terminate at the SOC on the opposite side of the brainstem or project to the lateral lemniscus. The remaining 25% of the fibers follow the pathway on the ipsilateral side of the brainstem and terminate at the SOC or the lateral lemniscus (Pickles, 1988). The SOC consists of a cluster of nuclei including the lateral superior olive (LSO), the medial superior olive (MSO), and the ventral nucleus of the trapezoid body (VNTB). The SOC is also the site at which afferent auditory neurons connect with facial nerve, which innervate the stapedius muscle in the middle ear. When a very intense sound traveling via the vestibulocochlear nerve arrives at the cochlear nucleus, the signal is sent to SOC via several different pathways (including ipsilateral and contralateral SOC) to the facial nerve nucleus. From the nucleus the signal travels via the facial nerve to the stapedius muscle, which contracts. Contraction of the stapedius muscle pulls the stapes posteriorly increasing stiffness of the ossicles and tympanic membrane and decreasing the effective level of transmission for loud low frequency sounds (Deutsch and Richards, 1979; Moller, 1965; Stach and Jerger, 1990; Wilber, 1976). In addition, VNTB projects to many cochlear nuclei and forms efferent medial olivocochlear bundle (MOCB) innervating ipsilateral and contralateral OHCs (Guinan, 2006; Warr and Beck, 1996) decreasing gain of the cochlear amplifier.
The cochlear nucleus, which spans the pons and medulla, is the first processing center and relay station of the ascending auditory pathways. The two other major relay stations in the brainstem are the superior olivary complex in the pons and the inferior colliculus in the midbrain. From the cochlear nucleus, the nerve fibers project to the superior olivary complex (SOC) or directly to the lateral lemniscus. Approximately 75% of the ascending CANS fibers leaving the cochlear nucleus cross over to the contralateral side of the brain to terminate at the SOC on the opposite side of the brainstem or project to the lateral lemniscus. The remaining 25% of the fibers follow the pathway on the ipsilateral side of the brainstem and terminate at the SOC or the lateral lemniscus (Pickles, 1988). The SOC consists of a cluster of nuclei including the lateral superior olive (LSO), the medial superior olive (MSO), and the ventral nucleus of the trapezoid body (VNTB). The SOC is also the site at which afferent auditory neurons connect with facial nerve, which innervate the stapedius muscle in the middle ear. When a very intense sound traveling via the vestibulocochlear nerve arrives at the cochlear nucleus, the signal is sent to SOC via several different pathways (including ipsilateral and contralateral SOC) to the facial nerve nucleus. From the nucleus the signal travels via the facial nerve to the stapedius muscle, which contracts. Contraction of the stapedius muscle pulls the stapes posteriorly increasing stiffness of the ossicles and tympanic membrane and decreasing the effective level of transmission for loud low frequency sounds (Deutsch and Richards, 1979; Moller, 1965; Stach and Jerger, 1990; Wilber, 1976). In addition, VNTB projects to many cochlear nuclei and forms efferent medial olivocochlear bundle (MOCB) innervating ipsilateral and contralateral OHCs (Guinan, 2006; Warr and Beck, 1996) decreasing gain of the cochlear amplifier.
The organization of superior olivary complex and the connecting neural fibers are shown in Figure 8-21. Ascending projections from both cochlear nuclei and both superior olivary complexes travel via the largest fiber tract in the CANS, the lateral lemniscus, to the inferior colliculi (one colliculus on each side), located on the posterior surface of the midbrain. Similar to the decussation seen at earlier levels in the brainstem, the two inferior colliculi are connected by fibers that allow crossover of signals from one side of the brainstem to the other. The connections between the two sides of the brainstem, from the SOC to the inferior colliculi, are important for directional hearing. From the inferior colliculi, all fibers ascend to the medial geniculate body in the thalamus. The thalamus is located immediately above the midbrain and it directs all sensory information (except smell) to the appropriate area of the cerebrum. The cerebellum, or “little brain,” is primarily responsible for coordinating motor commands with sensory inputs in order to control movement and communicates with the brainstem, spinal cord and cortex. Figure 8-20. Ascending pathway of the central auditory nervous system. Different parts of the auditory pathway are color coded
Figure 8-20. Ascending pathway of the central auditory nervous system. Different parts of the auditory pathway are color coded
 The cerebral hemispheres make up the largest portion of the forebrain. There are a number of connections between the medial geniculate body and the cerebral hemispheres, but the main ascending auditory pathway travels from the medial geniculate nucleus to the ipsilateral transverse temporal gyri (Heschl’s gyrus) of the cerebrum and then to auditory association areas in other areas of the brain.
The cerebral hemispheres make up the largest portion of the forebrain. There are a number of connections between the medial geniculate body and the cerebral hemispheres, but the main ascending auditory pathway travels from the medial geniculate nucleus to the ipsilateral transverse temporal gyri (Heschl’s gyrus) of the cerebrum and then to auditory association areas in other areas of the brain.
The outermost segment of the cerebrum is called the cerebral cortex; commonly referred to as the “gray matter.” The cortex is 2-6 mm (0.08 to 0.23 in) thick and is made up of the “gray-looking” nerve cell bodies. It is supported from underneath by the “white matter.” which consists of the myelinated nerve fibers (axons) connecting various gray matter areas of the brain to each other. The surface of the cortex contains numerous peaks (gyri) and valleys (sulci) that serve to increase the overall area of the cortex. Extremely deep sulci are called fissures.
The deepest fissure of the brain, the longitudinal fissure divides the cerebrum into two cerebral hemispheres. Cerebral hemispheres are only connected by a narrow structure called the corpus callosum, which is the only communication link between the hemispheres. Each hemisphere is divided into four basic anatomical areas called lobes. They are called the frontal lobe, temporal, parietal, and occipital lobes. The frontal lobe takes up 1/3 of the cortex and is associated with executive functions such as the planning and initiation of motor actions. The parietal lobe is the primary reception area for somatic sensory data, while the occipital lobe is the main visual processing center of the brain (Seikel, King and Drumright, 2000). The main site of auditory and receptive language (Wernicke’s area) processing centers is the temporal lobe.
The four lobes are additionally divided into smaller functional areas based on the type and organization of neurons occupying these areas. These areas are called Brodmann areas and numbered from 1 to 48. Many of them have also been found to be responsible for specific cortical activities and so are labeled by these activities. For example, auditory activity in the cortex has been found to be concentrated in Brodmann areas 41 and 42, which are called the primary auditory cortex, and in area 22, called the secondary auditory cortex. Both these regions are located in the posterior (back) part of the superior temporal gyrus and descend into the lateral sulcus (Sylvain fissure) as the transverse temporal gyri, known also as the Heschl’s gyri. A schematic view of various part of the cortex with a map of the Brodmann areas is shown in Figure 8-22. Figure 8-22. Sagittal view of the cerebral cortex and the Brodmann areas
Figure 8-22. Sagittal view of the cerebral cortex and the Brodmann areas
The cortex, and thus the auditory cortex, is organized in six neural layers numbered from I to VI (Emanuel and Letowski, 2009). Auditory information arriving at the thalamus is further relayed to nonpyramidal neurons located in layer IV of the primary auditory cortex. Layers V and VI have efferent connections to the medial geniculate nucleus and the inferior colliculus, respectively. Other layers are involved in motor function (layer II and III) and have connections to other parts of the brain. Through theses connections all information entering the brain creates a synergistic perceptual image of the surrounding environment together with corresponding emotional state created by this image.
|
181 videos|346 docs
|

video lectures
,practice quizzes
,Free
,study material
,mock tests for examination
,Extra Questions
,Objective type Questions
,Semester Notes
,Hearing System | Zoology Optional Notes for UPSC
,Hearing System | Zoology Optional Notes for UPSC
,Summary
,ppt
,past year papers
,Viva Questions
,MCQs
,Important questions
,shortcuts and tricks
,Sample Paper
,Previous Year Questions with Solutions
,Hearing System | Zoology Optional Notes for UPSC
,Exam
;






Hearing System Free PDF Download
Importance of Hearing System
Hearing System Notes
Hearing System UPSC Questions
Study Hearing System on the App
|
© EduRev
|
Education Revolution
|



|




