Type of Fossils and their Study Techniques | Botany Optional for UPSC PDF Download
| Table of contents |
|
| Method of Fossilization |
|
| Codaitales |
|
| Ginkogales-A living fossil |
|
| Male and Female gametophyte of Pinus |
|
| Life cycle of Gnetum and its angiospermic feature |
|
| Bennettitales |
|
| Pentoxylon |
|

Method of Fossilization
The term "fossil" encompasses any evidence of past life, which can include the preserved remains of organisms like plants, shells, teeth, or bones. Fossils can also capture traces of an organism's activity. Various processes lead to the formation of fossils, with a common one being the death of a plant or animal in a water-rich environment, followed by burial in mud and silt. Soft tissues decompose rapidly, leaving behind the durable bones or shells. Over time, sediment accumulates and hardens into rock. It's during erosion processes that these preserved remnants in stone are uncovered. Fossils, which are the remains of ancient plant and animal life, are typically discovered within sedimentary rocks, with shale, limestone, and sandstone being common hosts for fossils. Earth comprises three rock types: metamorphic, igneous, and sedimentary.
The history of earth can be described into different era which has been shown below: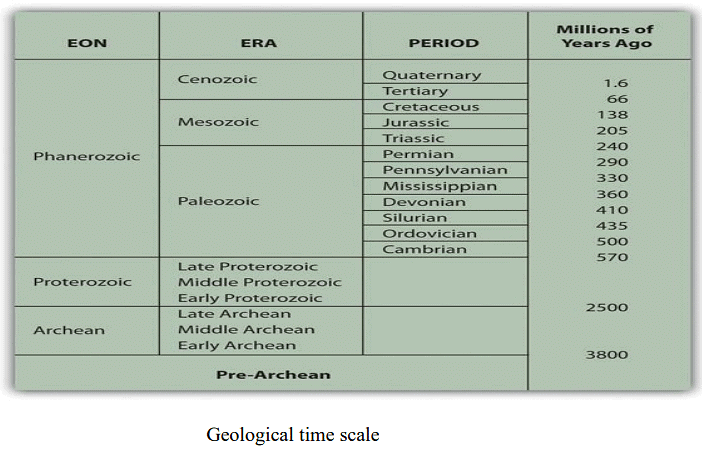
Animals which survived as major Biodiversity during different era has been shown below: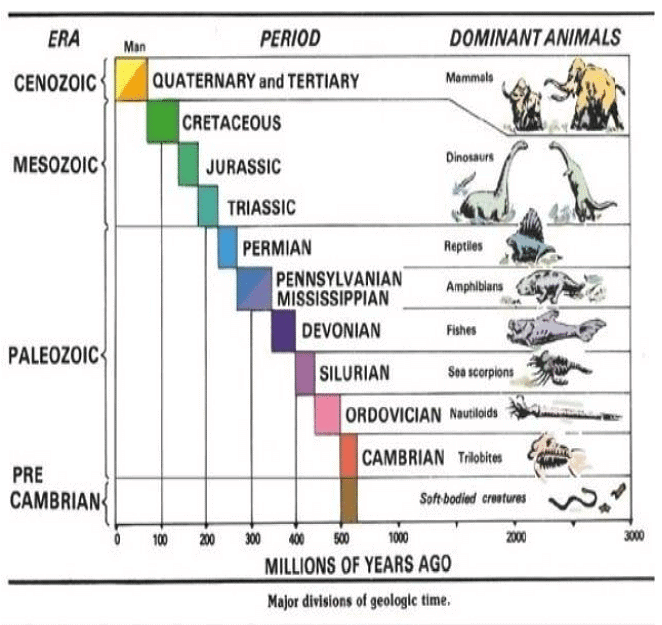 Plants which survived as major Biodiversity during different era has been shown below:
Plants which survived as major Biodiversity during different era has been shown below: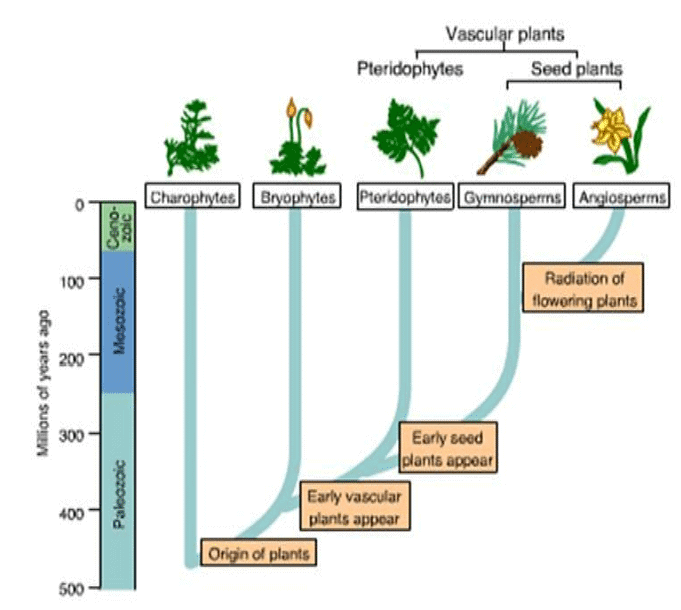
Exploring different eras of Earth reveals the appearance of distinct plants and animals during various periods. Earth has preserved these remnants as fossilized flora and fauna. The fossil record has been instrumental in tracing the origins and evolutionary history of diverse plants and animals.
Process of Fossilization
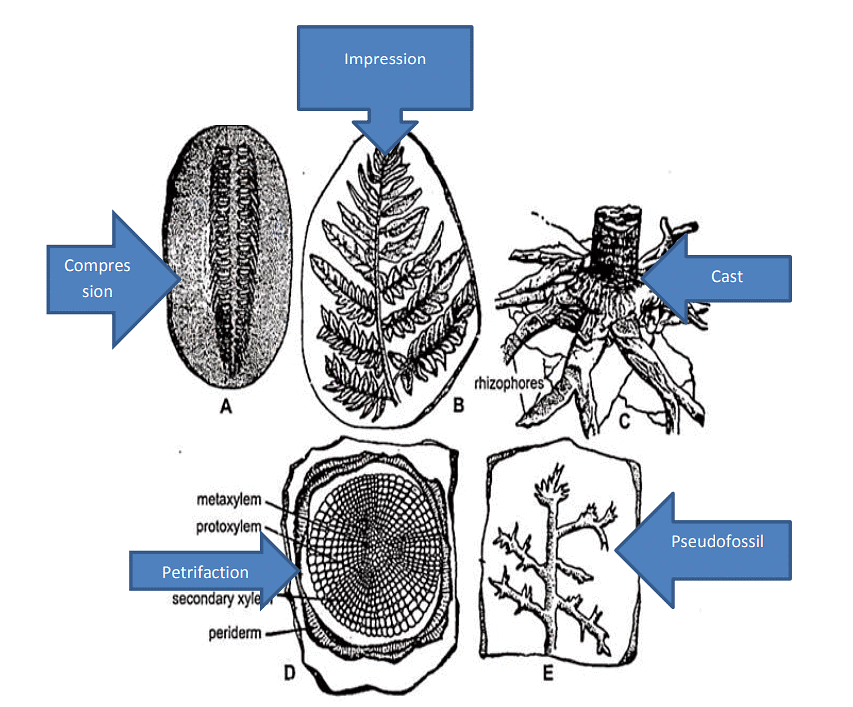
The five most often cited types of fossils are mold, cast, imprint, permineralization and trace fossils.
- Mold or Impression Fossil: This type of fossil forms when a plant or animal completely decays but leaves behind an imprint or hollow mold of itself.
- Cast Fossil: When an organism dies, its flesh decomposes, and its bones deteriorate due to chemical reactions. Minerals gradually enter the cavity, creating a mineralized replica of the organism left in the sediment, known as a cast.
- Imprint Fossil: Imprint fossils result from an organism's movement, leaving behind traces or tracks. These tracks are preserved when clay or silt dries slowly and gets covered by additional sediment. Imprint fossils can also occur when plants are covered by sediment.
- Permineralization Fossil: One of the common fossilization types is permineralization. This process involves the impregnation of the pores in plant materials, bones, and shells with minerals from the surrounding ground, lakes, or oceans. In some cases, wood fibers and cellulose dissolve, and minerals take their place. Two typical forms of permineralization are silicification and pyritization.
Fossils are of various types some of which has been described below:
- Petrified Fossils: These fossils result from the gradual replacement of plant parts by minerals like iron, pyrites, silicates, carbonates, and sulphates. These minerals infiltrate and deposit themselves molecule by molecule within the cells and tissues of the plant. To study the structural details, sections of these fossils can be prepared.
- Compression Fossils: Commonly found in sedimentary rock deposits, compression fossils are essentially impressions where a majority of the organic remains of a plant are preserved. The plant or plant part becomes buried, and sediments continue to accumulate over it.
- Impression Fossils: These fossils consist of impressions of plants or plant parts left on sediments. They are valuable for studying the external features of various plant parts and the patterns of venation in leaves.
- Trace Fossils: These fossils provide insight into the activities of organisms. For instance, an animal leaves behind a footprint when it steps on sand.
- Preserved Remains or Amber Fossils: Some organisms are exceptionally well-preserved, often near their original states. These fossils are referred to as preserved remains. For example, an insect may become trapped in the sticky resin of a tree and eventually die. As more resin covers it, the insect is sealed inside, and over time, it hardens into amber.
Codaitales
Distinguishing Features of Cordaitales
- Tree Structure: Cordaitales were characterized by tall trees with slender trunks and a crown of multiple well-developed branches.
- Temporal Range: These plants existed from the Devonian to Permian periods of the Palaeozoic era.
- Leaf Characteristics: Cordaitales had simple leaves that were spirally arranged. These leaves could be strap-shaped, grass-like, or paddle-like in appearance.
- Leaf Size: The leaves of Cordaitales could grow to lengths of up to 1 meter or even longer. They exhibited parallel venation.
- Wood Structure: These plants had a limited primary wood. In mature stems, the secondary wood was predominantly pycnoxylic.
- Reproductive Structures: Cordaitales featured compound unisexual cones.
- Cone Structure: Each compound cone had a central axis with bracts that subtended secondary fertile shoots. These secondary shoots possessed both fertile and sterile appendages.
- Female Cones: The megastrobili (female cones) had sterile appendages below and ovule-bearing fertile appendages above.
- Ovules: Typically, one to four ovules were found on each female fertile appendage.
- Male Cones: The microstrobili (male cones) had sterile appendages below and fertile appendages containing pollen sacs above.
- Pollen Sacs: Each male fertile appendage typically carried four to six terminal pollen sacs.
- Sperm Production: Although actual sperm cells haven't been reported, the presence of pollen chambers suggests that motile sperm might have been produced in Cordaitales.
Features of Cordaitales of Cordaitales
Morphological Features:
- The Cordaitales plant had a tall and slender structure.
- Near the top of the plant, there was a crown of branches.
- The leaves of Cordaitales were large, simple, and pendulous (hanging down). It's worth noting that leaves with these characteristics are not found in any living gymnosperms.
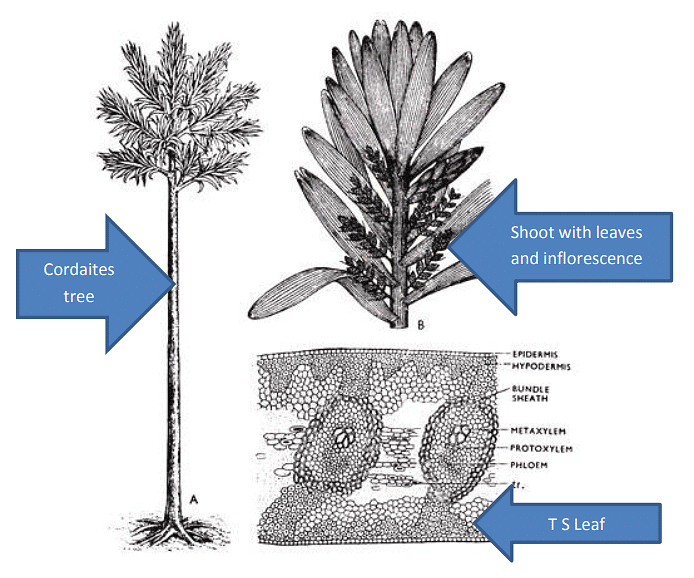
Anatomical Features:
- The internal structure of the stem in Cordaites exhibited a combination of characteristics seen in both Cycas and conifers.
- The pith in Cordaites resembled that of cycads, being discoid in form and arranged in plate-like structures at intervals.
- The vascular cylinder, on the other hand, resembled that of conifers. It consisted of thick collateral (having xylem and phloem located side by side) endarch (xylem matures centripetally) primary bundles. In some instances, these bundles appeared to be mesarch (xylem matures both centripetally and centrifugally), similar to Mesoxylon.
- Cordaitales displayed substantial normal secondary growth, indicating the development of additional vascular tissue over time.
- The secondary xylem in Cordaitales was composed of living pitted tracheids with pits on their radial walls.
- Interestingly, many plants from the Carboniferous period, including those belonging to Cycadofilicales, Cordaitales, and several fossil pteridophytes, exhibited similar anatomical features.
- While these plants had significant secondary growth in their stems, they did not form annual rings in their growth patterns.
- The stem anatomy of Cordaitales demonstrated variations in the arrangement of vascular bundles, ranging from exarch (xylem matures outward) to end-arch (xylem matures inward) collateral primary bundles. This diversity in vascular bundle arrangement is similar to that found in Cycadofilicales and gymnosperms, particularly conifers.

Reproductive Structures of Cordaitales
- The plants were monoecious or dioecious but the strobili were always monosporous and never bisporangiate like Cycadeoideales (Bennettitales) or living Gnetales. Cordaitanthus was the reproductive shoot of Cordaites itself. It was not definitely organized to form a cone but it was more or less a strobilus.
- The male and female strobili were borne separately in lateral sides of short simple axis. The strobili were enclosed when young by bracts so outwardly they could not be distinguished unless sections were cut and examined. Both were usually of small size.
Strobilus of Cordaitales
Male Strobilus
- The male strobilus of Cordaitales had a thick central axis with spirally arranged bracts, each bearing stamens.
- The stamens could either be solitary or grouped near the apex, with sterile bracts present in between.
- Each stamen had a unique structure, with a cluster of three to six terminal sporangia.
- Some stamens had synangia and individual sporangia at their tips.
- The male strobilus of Cordaitales resembled the male reproductive structure of Ginkgo, although there was a difference in the placement of sporangia, which were terminal in Cordaitales and pendant in Ginkgo.
Female Strobilus
- The young female strobilus was enclosed by bracts and had a thick conical axis.
- Dwarf shoots were present in the axils of some bracts, each having its own minute bracts.
- At the tip of each dwarf shoot, there was a terminal ovule.
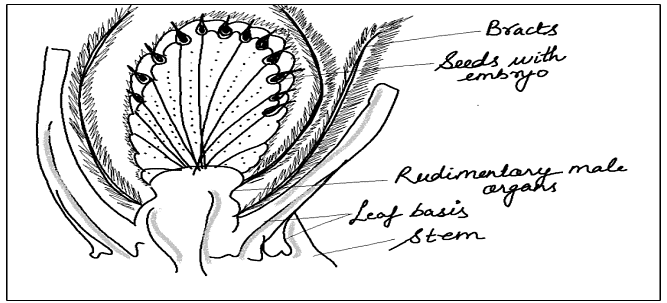
Ovule of Cordaitales
- In Cordaitales, the ovule had a distinct structure. The nucellus was separate from the integument, forming a micropyle.
- A prominent nucellar beak and a large pollen chamber were present at the base of the micropyle.
- The ovule was surrounded by two coats: an outer thick and fleshy one and an inner delicate one in young ovules, which became hard in older ones.
- These coats were separate below but more or less fused above.
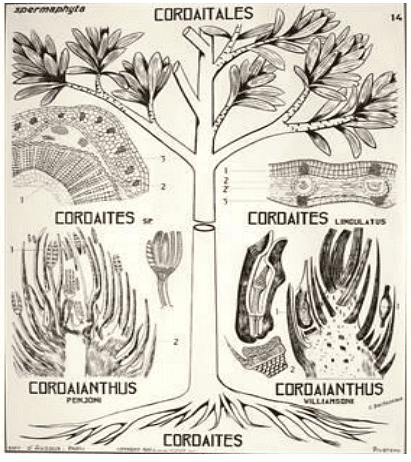
Evolutionary Affinities
- Cordaitales and Cycadofilicales coexisted during the Paleozoic era, particularly in the Carboniferous period, and both disappeared around the Permian.
- The exact ancestral relationship between the two is not definitive, but they share numerous primitive features, such as the presence of free nucellus, the form of integument with vascular bundles, the shape of the pollen chamber, and the absence of an embryo.
- It is possible that they both evolved from a common ancestor, possibly a heterosporous pteridophyte from the Devonian period.
- Cordaitales made advancements in stem anatomy and reproductive complexity, differentiating itself from Cycadofilicales. It is considered a more advanced group.
- Cordaitales might have given rise to Ginkgoales and conifers before disappearing during the Permian period, sharing resemblances in stem anatomy, ovule structure, and the presence of sperms with these groups.
Ginkogales-A living fossil
Salient features of Ginkgo
The plant body of Ginkgo biloba is sporophytic, and the sporophyte resembles several conifers in general habit. The branches are dimorphic i.e. bear long shoots which are of unlimited growth with scattered leaves and dwarf shoots which are short branches of limited growth.
The foliage leaves, present on the long shoots, are deeply lobed while those on the dwarf shoots are not so deeply lobed and sometimes more or less entire. In general, the leaves are bilobed, and hence the name ‘biloba’ was suggested to the species by Linnaeus (1771). The leaves of the long shoot and seedlings are deeply bilobed while that of the dwarf shoot are not so deeply lobed and may be sinuate or even entire. They may be pale yellow, golden yellow or dark green in colour. A typical dichotomous type of venation is present in the leaves.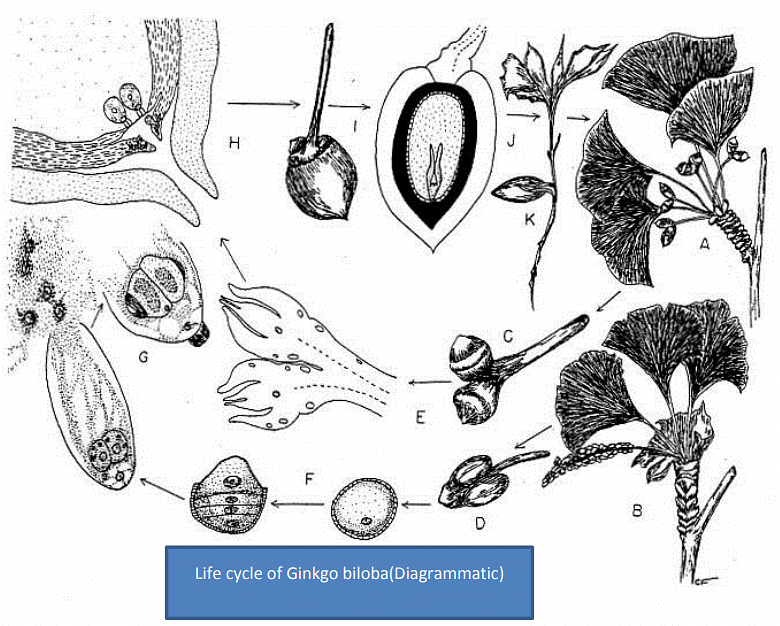
Anatomy of Ginkgo
Root
- In cross-section, Ginkgo roots have a roughly circular shape.
- Mature roots are encased by a layer of phellogen or suberized cells in the cortex.
- Young roots contain a significant portion of multilayered, thin-walled cortex with tannin-filled cells and calcium oxalate crystals.
- Prominent mucilage canals are observed in the roots.
- Young roots exhibit distinct layers of endodermis and a single-layered pericycle, but mature roots lack such differentiation.
- Diarch condition is evident in young roots, and the xylem is exarch.
- The xylem remains separated by phloem strands, and triarch conditions may also be observed in some roots.
Stem
- Young stems (long shoots) have a roughly circular shape and are surrounded by a single-layered, thick, circularized epidermis composed of brick-shaped cells.
- Older stems develop a periderm that replaces the epidermis.
- The stem's cortex contains mucilaginous canals, sphaeraphides, and numerous tannin-filled cells.
- Cortex is relatively less extensive in long shoots compared to dwarf shoots.
- Endodermis and pericycle are not distinctly marked in long shoots.
- Very young stems have several conjoint, collateral, open, and endarch vascular bundles arranged in a ring.
- As secondary growth begins, the stem's vascular cylinder transforms into an endarch siphonostele, with no parenchyma in the wood except for uniseriate medullary rays.
- Protoxylem is characterized by spiral thickenings, and bordered pits are present on the radial walls of metaxylem tracheids.
- The phloem consists of sieve tubes and phloem parenchyma.
- In the center of long shoots, a narrow pith containing mucilage canals and sphaeraphides is found, while dwarf shoots have a relatively larger pith.
- Cork cambium contributes to secondary growth, producing periderm that replaces the epidermis. Mucilage canals are absent.
- A single ring of cambium remains active throughout and generates secondary phloem toward the outer side and secondary xylem toward the inner side.
- One or two rows of circular pits with clear tori are present on the radial walls of the tracheids. It's important to note that these pits differ from those in the primary wood, and there are also trabeculae of Sanio, which cross the tracheids' lumens.

Leaf
- Ginkgo leaves have upper and lower epidermal layers, both thickly cuticularized.
- Stomata of the haplocheilic type are restricted to the lower epidermis.
- Mesophyll is located between the two epidermal layers and lacks a clear differentiation into palisade and spongy parenchyma, except in leaves of long shoots.
- The mesophyll region contains mucilage canals or secretory canals and a few tannin-filled cells.
Petiole
- The petiole is covered by a layer of thickly cuticularized epidermis, featuring stomata.
- Several hypodermal layers are found beneath the epidermis.
- Mucilage canals, tannin-filled cells, and sphaeraphides are irregularly distributed in the petiole's cortex.
- A pair of endarch vascular bundles supplies the petiole, enclosed by a sclerenchymatous bundle sheath.
- Protoxylem in the petiole has spiral thickenings, and cambium is visible with uniseriate rays present in the xylem, which are continuous with those of the phloem.
Reproductive Structures of Ginkgo
- Ginkgo biloba is a dioecious plant, which means that male and female plants exist, but they are challenging to distinguish when young. The reproductive structures of Ginkgo are considered among the most primitive among living seed plants, similar to some Cycadales.
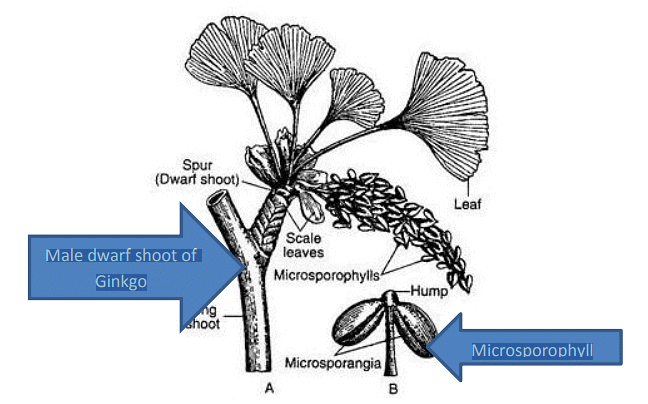
(i) Male Strobilus:
- Male or microsporangiate fructifications develop in clusters resembling catkins on the dwarf shoots of male Ginkgo trees.
- Each male strobilus consists of multiple microsporophylls loosely arranged on a central axis.
- Within each microsporophyll, there are typically two pendant microsporangia, although sometimes more may be present.
- Each sporangium is a tubular structure surrounded by several layers of tissue. The outermost layer differentiates into a tapetum.
- The sporogenous cells within the sporangium undergo reduction division, resulting in the formation of numerous haploid microspores.
(ii) Development of Microsporangium:
- The development of the microsporangium follows the eusporangiate type, where a single archesporial cell divides through a periclinal wall, forming a primary wall cell and a primary sporogenous cell.
- The primary wall cell contributes to the sporangium's wall, while the primary sporogenous cell develops into the sporogenous tissue.
- Dehiscence of the sporangium occurs through a longitudinal slit.
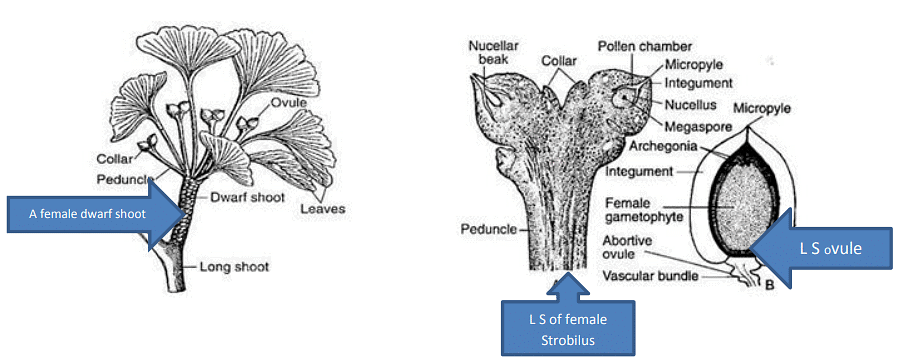
(iii) Female Strobilus:
- Female strobili are found in groups in the axils of leaves or scaly leaves on the dwarf shoots.
- These structures are highly reduced and each bears a long stalk or peduncle, which typically bifurcates at the apex, with one of the bifurcations often bearing a single ovule.
- Usually, one of the two sessile ovules present aborts at an early stage.
- Each ovule has a cup-like structure called a ring or collar at its base.
- Unlike the bilobed leaves surrounding them, the leaves around the ovules do not exhibit this characteristic.
- The development of the ovule, the process of megasporogenesis, and the structure of the mature ovule in Ginkgo are similar to those in Cycas.
- The ovule has a thick integument consisting of three layers: an outer fleshy layer, a middle stony layer, and an inner fleshy layer.
- A notable difference from Cycas is the lack of a distinct vascular supply in Ginkgo's integuments.
- Each ovule contains a prominent nucellus, and the free apex of the nucellus breaks down to form a pollen chamber, creating the nucellar beak.
- Within the nucellus tissue, a functional spore mother cell becomes prominent, developing into a tetrad of spores. However, only the innermost megaspore remains functional and matures into the female gametophyte.
- The vascular supply of the ovule is not well-developed, with two vascular strands entering the inner fleshy layer and extending up to the free part of the nucellus without branching, while the outer fleshy layer lacks significant vascular supply.
Gametophytes in Ginkgo
(i) Male Gametophyte
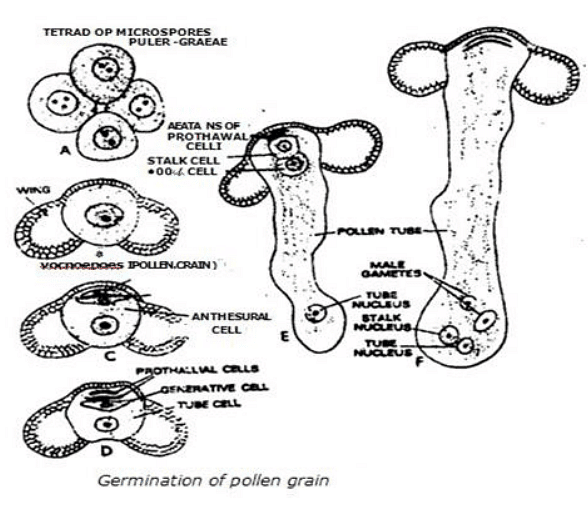
- The development of the male gametophyte in Ginkgo biloba shares similarities with that of Pinus. It begins to develop within the sporangium, and pollen germination initiates with the swelling of the tube cell. The tube cell becomes highly elaborate, consisting of a proximal unbranched tube and an extensively branched haustorial system.
- Each microspore is the first cell of the male gametophyte and has a rounded structure with thin intine and thick exine layers. It contains a centrally located nucleus with one or two nucleoli, surrounded by dense cytoplasm. Each microspore also has a region called the pore, where the exine does not cover the intine.
- The first mitotic division in the microspore results in two unequal cells, known as the 1st prothallial cell and the inner cell. The 1st prothallial cell is smaller than the inner cell, and the inner cell further divides to form a 2nd prothallial cell and an antheridial initial. During this time, the 1st prothallial cell begins to degenerate. The antheridial initial eventually divides to produce a generative cell near the 2nd prothallial cell and a tube cell.
- This stage represents the 4-celled male gametophyte, and at this point, the microspores are dispersed by the wind. Pollination in Ginkgo biloba occurs in April, and further development of the male gametophyte takes place in the pollen chamber. Here, the generative cell of the 4-celled male gametophyte divides into a stalk cell, close to the 2nd prothallial cell, and a body cell.
- The intine protrudes near the pore, functioning as a pollen tube, which swells and advances toward the archegonia by extending into the nucellus. Just before fertilization, the body cell undergoes division, and the resulting cells form sperms or male gametes. These sperms are multi-flagellated antherozoids.
(ii) Female Gametophyte
- Out of the four megaspores produced from the megaspore mother cell, only the lowermost remains functional, while the other three degenerate. The nucleus of the functional megaspore migrates toward the micropylar end and undergoes division, followed by multiple free-nuclear divisions, resulting in the formation of numerous free nuclei. A large vacuole is present in the megaspore at this stage.
- Wall formation commences, progressing from the periphery toward the center, with the formation of anticlinal walls followed by vertical walls. Each cell generally contains one nucleus, although some cells may have 2-3 nuclei. Due to faster cell divisions at the micropylar end, a pole-like structure called a tent-pole develops.
- The female gametophyte is rich in chlorophyll and possesses abundant chloroplasts.
Development of Female Gametophyte
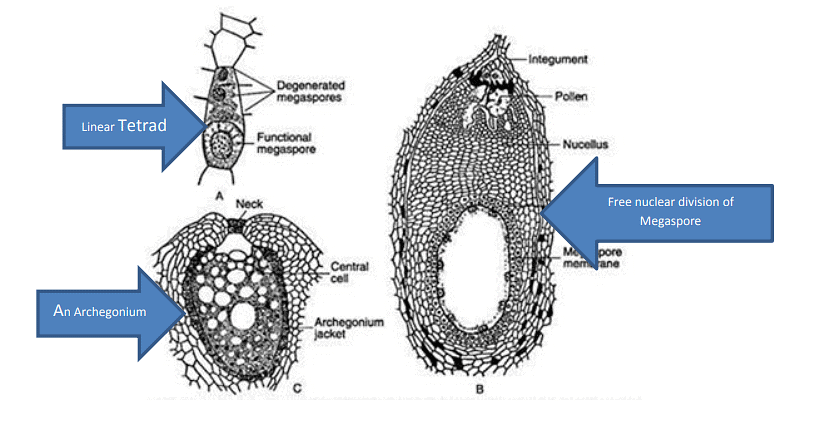
The development of archegonium starts from the cells towards the micropylar end of the female gametophyte. Each archegonium possesses a short neck made up of only four cells and a small venter having a central cell. The central cell later on forms a ventral canal cell and an egg cell. According to Bierhorst (1971) only two cells are present in the archegonial neck.
Embryogeny in Ginkgo
- The zygote becomes enlarged and starts dividing by many free-nuclear divisions just after fertilization. The nuclei are distributed irregularly . The cell wall formation starts just after a few divisions of the nucleus and soon a pro-embryo is differentiated . Three types of cells are clearly differentiated in the pro-embryo.
- The cells towards the chalazal end are smaller in size with dense cytoplasm, followed by another zone of comparatively larger cells, which do not possess such a dense cytoplasm. Elongated cells are present in the uppermost zone. There is no differentiation of suspensor in the embryo.
- The development of the cotyledon starts from the lowermost zone soon after the differentiation of these zones . Roots, stem and cotyledons are soon differentiated. Generally, five leaves are present in the mature embryo. Out of these five leaves the first two are decussate with cotyledons while the remaining leaves are irregular in arrangement in embryo and seedling.
Germination of Seed
The germination of the seed is of hypogeal type and quite similar to that of Cycas. A strong tap root develops soon and the seedling bears many bilobed leaves, which is characteristic feature of Ginkgo biloba. Soon, a mature plant, with many more leaves, develops.
Male and Female gametophyte of Pinus
Male gametophyte of Pinus
The male gametophyte of Pinus, also known as the pine tree, undergoes a series of developmental stages:
- Microsporophyll: This is the structure that contains the microspores. The microsporangium, which produces several microspores, is sessile and surrounded by a single layer of cells.
- Microspore Formation: Each microsporangium generates multiple microspores. Each microspore is enclosed by an inner intine and an outer exine layer. These microspores are winged.
- Germination of Microspore: The microspore serves as the initial cell of the male gametophyte and begins to germinate within the microsporangium.
- Early Division: The microspore nucleus undergoes division, resulting in the formation of a very small first Prothallial cell and a large cell.
- Further Division: The large cell divides, giving rise to a second Prothallial cell and an antheridial cell. At this stage, the microspore is shed and falls near the female gametophyte.
- Development Inside Female Gametophyte: Once inside the female gametophyte, the two Prothallial cells degenerate, and the antheridial cell divides again to produce a small generative cell and a large tube cell (also known as the tube nucleus).
- Pollen Tube Formation: The tube nucleus is responsible for regulating the growth of the pollen tube, which extends in close proximity to the nucellus of the female gametophyte.
- Attachment to Ovule: A mature ovule (megasporangium) secretes a mucilaginous substance from the micropyle. Numerous pollen grains become attached to this substance.
- Pollen Tube Penetration: As the mucilage dries up and reaches the apex of the nucellus, some of the pollen grains are drawn inside. The pollen tube, formed as a proliferation of the intine from the microspore, penetrates the nucellus.
- Generative Cell Division: Within the pollen tube, the generative cell divides to produce a sterile stalk cell and a fertile body cell.
- Formation of Male Gametes: The body cell further divides to create two non-motile male gametes (sperms). This marks the completion of the male gametophyte development in Pinus.
Female gametophyte of Pinus
The development of the female gametophyte (archegonia) in Pinus, commonly known as pine trees, involves the following steps:
- Megasporophyll Structure: The female cone of Pinus consists of megasporophylls, each composed of a large ovuliferous scale and a smaller bract scale. These scales are separate from each other.
- Ovules on Ovuliferous Scale: Each ovuliferous scale bears two anatropous ovules or megasporangia. These ovules are positioned on the upper surface of the ovuliferous scale toward its base.
- Megasporangium Structure: Each megasporangium consists of two main parts:
- Nucellus: This is a substantial tissue within the megasporangium.
- Integument: The integument covers the nucellus. At the basal portion of the integument, it is fused with the nucellus, and at the top, it opens to form a micropyle, which is an opening in the integument.
- Megaspore Mother Cell: Within the nucellus, a single megaspore mother cell is differentiated. This megaspore mother cell undergoes meiotic division to produce a row of four megaspores.
- Functional Megaspore: Among the four megaspores formed, only the lowermost one remains functional, while the others degenerate. This functional megaspore increases in size and initiates the development of the female gametophyte, which includes the formation of archegonia.
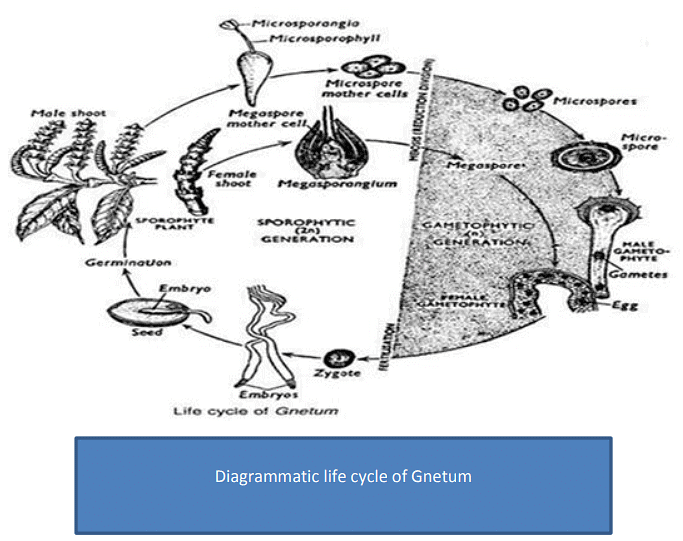
Life cycle of Gnetum and its angiospermic feature
Gnetum (35 species) inhabits a variety of humid, tropical lowland, riparian, and swamp rainforests of South East Asia, Papua New Guinea, Fiji, the Americas, and Africa . Gnetum bears remarkably angiosperm-like leaves, consisting of a broad, entire-margined lamina with pinnatereticulate venation and multiple vein orders The majority of Gnetum species are large woody climbers, producing xylem from multiple cambia. Some Gnetum vines ascend high into the canopies of dense riparian vegetation and lowland forest trees; other species occur as low scramblers in open, fire-burnt pastures and disturbed forest edges. Two Gnetum taxa (i.e., G. gnemon, widespread in the Indo-Pacific; G. costatum, from eastern Papua New Guinea and the Solomon Islands), that form sun-exposed medium-sized subcanopy trees in lowland rainforest and riverine gallery forests (typically 7 to 15 m, as high as 20 m) possess some peculiar lianalike features. In addition to the occasional production of scandent branches, older trees of G. gnemon develop additional anomalous cambia in the bark that are akin to multiple cambia of lianoid Gnetum species . Interestingly, molecular phylogenetic analyses indicate that treeforming Gnetum species are well nested among Gnetum climbers, suggesting that arborescence is secondarily derived.
Habit of Gnetum
Majority of the Gnetum species are climbers except a few shrubs and trees.The branch is of limited growth ,unbranched and bears decussate foliage leaves.The leaves are oval in shape having entire margin showing reticulate venation.
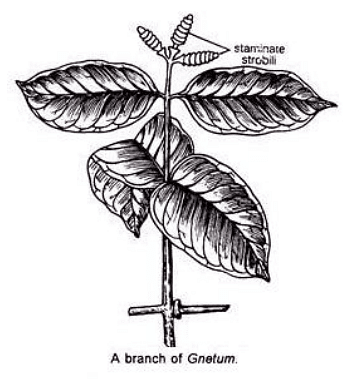
Anatomy of Gnetum
Root
- The young root of Gnetum has multiple layers of starch-filled parenchymatous cortex, with large polygonal cells.
- An endodermal layer is distinguishable, and it contains Casparian strips in its cells.
- The endodermis is followed by a 4-6 layered pericycle.
- Gnetum roots are diarch (having two protoxylem groups) and exarch (with xylem located external to phloem).
- In young roots, a small amount of primary xylem is visible, but it becomes indistinguishable after secondary growth.
- Secondary growth in roots is of a normal type and includes tracheids, vessels, and xylem parenchyma. Tracheids have uniseriate bordered pits with bars of Sanio, and vessels have simple or small multiseriate bordered pits.
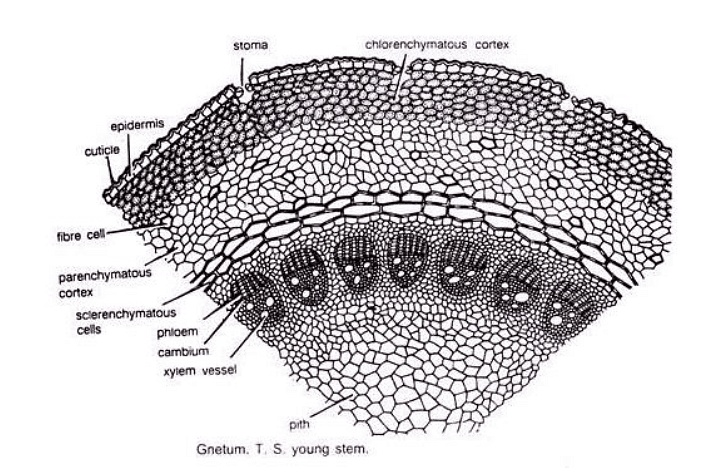
Young Stem
- The transverse section of the young stem is roughly circular and resembles a typical dicotyledonous stem.
- It is surrounded by a single-layered epidermis with thick, circularized walls made of rectangular cells.
- The cortex consists of an outer chlorenchymatous region (photosynthetic tissue), a middle parenchymatous region, and an inner sclerenchymatous region.
- Several conjoint, collateral, open, and endarch vascular bundles are arranged in a ring in the young stem.
- Xylem in the stem includes tracheids and vessels, which is a feature more commonly associated with angiosperms. Protoxylem elements are spiral or annular, and metaxylem has circular-bordered pits.
- Phloem consists of sieve cells and phloem parenchyma.
Old Stem
- In older stems of Gnetum, secondary growth is observed.
- In Gnetum gnemon, secondary growth is normal, resembling that of dicotyledonous plants.
- However, in many Gnetum species (e.g., G. ula, G. africanum), an anomalous type of secondary growth occurs.
- The primary cambium is short-lived, and a series of secondary cambia develop in the cortex, forming successive rings of secondary tissue.
- This results in eccentric rings of secondary xylem and phloem in the wood, representing an abnormal growth pattern.
- Secondary wood in old stems consists of tracheids and vessels. Tracheids have bordered pits on their radial walls, while vessels have simple pits.
Reproduction of Gnetum
Gnetum is dioecious. The reproductive organs are organised into well-developed cones or strobili. These cones are organised into inflorescences, generally of panicle type. Sometimes the cones are terminal in position.
Male Cone and Male Flower
- The male flowers are arranged in definite rings above each collar on the nodes of the axis of male cone. The number of rings varies between 3-6. The male flowers in the rings are arranged alternately. There is a ring of abortive ovules or imperfect female flowers above the rings of male flowers.
- Each male flower contains two coherent bracts which form the perianth. Two unilocular anthers remain attached on a short stalk enclosed within the perianth from which it comes out during maturity to dehisce.
Development of Male Flower
- In very young cones, certain cells below each collar become meristematic. They divide repeatedly and form a small hump-like outgrowth. Certain cells on the upper side of this annular outgrowth start to differentiate into the initials of the ovules. They develop into abortive ovules which form the uppermost ring. The cells of the lower side of this annular outgrowth form the primordium of male flower.
- A central cushion of cells develops by the repeated divisions in the male flower primordium. This cushion gets surrounded by a circular sheath called perianth. The sheath-like perianth encloses the central cushion-like mass only partially. With the development of a depression or notch in the central mass two lobes differentiate and later on develop into two anther lobes.
- With the help of many divisions the basal portion of this central mass of cells starts to differentiate into a stalk. This stalk elongates and pushes the anther lobes towards the outer side. Each anther lobe remains surrounded by an epidermal layer and a few wall layers which enclose a microsporangium.The innermost wall layer enclosing the sporogenous tissue is known as tapetum.
- The sporogenous cells become loose, contract round up and change into the spore mother cells. In the process of microspore formation the tapetum and two wall layers are used for the developing microspores. The spore mother cells undergo meiosis and ultimately the spore tetrads are formed.
Female Cone
The female cones resemble with the male cones except in some definite aspects. A single ring of 4-10 female flowers or ovules is present just above each collar . Only a few of the ovules develop into mature seeds In the young condition, there is hardly any external difference between female and male cones. All the ovules are of the same size when young but later on a few of them enlarge and develop into mature seeds. All the ovules never mature into seeds.Each ovule consists of a nucellus surrounded of three envelopes. The nucellus consists of central mass of cells. The inner envelope elongates beyond the middle envelope to form the micropylar tube or style. The nucellus contains the female gametophyte. There is no nucellar beak in the ovule of Gnetum.
Fertilization in Gnetum
- After the entry of pollen tube into the female gametophyte, whose lower end has become partly cellular, one or more nuclei (3-8) at its upper end also become delimited by cell-walls. These groups of cells are usually present in the neighbourhood of the pollen tube. Out of each such group, one or rarely two cells behave as egg cells, which often become surrounded by one or two layers of minute and degenerated cells.
- In all probability these inconspicuous cells provide nourishment to the egg and become used up at the time of fertilization. Only a male nucleus enters the egg, while its sheath is left outside. In cases where two eggs are present in the vicinity of the pollen-tube, double fertilization may result.
- Gnetum is remarkable among gymnosperms in the fact that a typical gymnospermic endosperm, which is formed prior to fertiliza-tion, is lacking here. Madhulata (1960) reports in G. gnemon that while eggs are being differentiated, the female gametophyte re-mains in a free nuclear condition, and only when zygotes are produced in the micropylar region of the ovule, the first walls are laid down in the chalazal region producing in an upward direction.
- Very rarely the cell-wall formation may take place simultaneously throughout the gametophyte. Sometimes walls may be laid down even when eggs are not differentiated within the gametophyte.
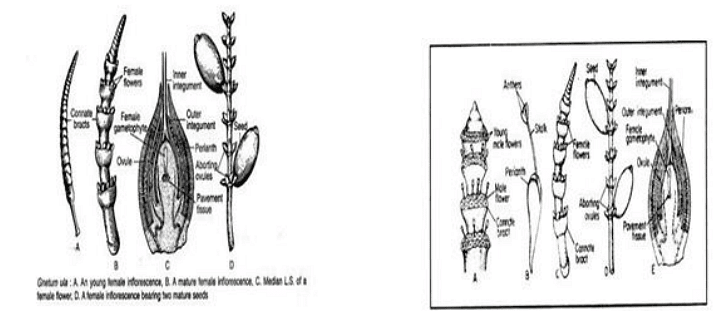
Angiospermic feature of Gnetum
A key position to Gnetum has been assigned by scientists while discussing the origin of angiosperms. Both Gnetales and angiosperms originated from a common stalk called “Hemiangiosperm”.
In a beautiful monograph on Gnetum, Maheshwari and Vasil (1961) are of the view that Gnetum possesses some strong angiospermic features” and hence they are close to angiosperms.
Some of the resemblances between Gnetum and angiosperms are mentioned below:
- General Habit: The sporophyte of many Gnetum species shares similarities in its general habit with angiosperms.
- Reticulate Venation: Gnetum leaves display reticulate venation, a characteristic commonly associated with angiosperms.
- Endarch Vascular Bundles: The presence of endarch vascular bundles in Gnetum is a feature shared with dicot angiosperms.
- Vessels in Xylem: Gnetum possesses vessels in its xylem, which is a characteristic typically associated with angiosperms.
- Shoot Apex Configuration: Both Gnetum and angiosperms exhibit clear tunica and corpus configurations in their shoot apices.
- Strobili Resemblance: The strobili (reproductive structures) of Gnetum bear a closer resemblance to those of angiosperms compared to other gymnosperms.
- Micropylar Tube: The micropylar tube of Gnetales performs functions similar to the style in angiosperms.
- Tetrasporic Development: Gnetum displays tetrasporic development of the female gametophyte, a feature that aligns it with angiosperms.
- Absence of Archegonia: The absence of archegonia in both Gnetum and angiosperms further underscores their similarities.
- Dicotyledonous Embryo: The dicotyledonous nature of the embryo in Gnetum makes it more closely related to dicotyledonous angiosperms.
- Companion Cells: Gnetum shares the presence of companion cells with angiosperms, which is a consistent anatomical feature.
- Double Fertilization: Both Gnetum and angiosperms undergo double fertilization, a unique reproductive feature not found in other gymnosperms.
- Production of Nectar: Some members of Gnetopsida, including Gnetum, produce nectar, a feature also found in angiosperms.
Bennettitales
The name "Bennettitales" is in honor of J.J. Bennett, an English botanist. The first specimen of Bennettitales was discovered in 1825, which was a fossilized trunk of the genus Bucklandia collected from Great Britain. In 1867, a silicified trunk of Cycadeoidea etrusca was found in an Etruscan tomb.
Williamson (1870) introduced the name "Williamsonia" for a combination of foliage and reproductive organs found in Williamsonia gigas. Professor Birbal Sahni (1932) discovered Williamsonia sewardiana from the Upper Gondwana beds of India.
Rich fossil flora of Bennettitales has been reported from the Rajmahal Hills of Bihar, India. Some of the reported members include Bucklandia sahnii, B. indica, Dictyzamites, Otozamites benghalensis, Cycadinocarpus rajmahalensis, Sahnioxylon rajmahalensis, Williamsonia indica, W. sahnii, and W. sewardiana.
Arnold (1948) classified Bennettitales into two families: Williamsoniaceae and Cycadeoideaceae, while Sporne (1965) further divided them into three families:
- Williamsoniaceae, including Williamsonia and Pterophyllum.
- Wielandiellaceae, including Wielandiella and Williamsoniella.
- Cycadeoideaceae, including Cycadeoidea (also known as Bennettites).
Distinguishing features of Bennettitales include:
- These extinct Mesozoic plants existed on Earth from the Triassic to the Cretaceous period.
- Bennettitales were so abundant during the Mesozoic era that this period is known as the 'Age of Cycads.'
- Fossils of Bennettitales are typically found as compressions or petrifactions.
- The stems of Bennettitales were stout or slender and had a wide pith.
- The stems grew very slowly and had manoxylic wood, characterized by a lack of compactness, a large pith, and a cortex mixed with a smaller amount of xylem tracheids. Parenchyma cells in the wood were filled with starch grains, but this wood was not durable.
- The leaves of Bennettitales were mostly pinnately compound, similar to living Cycads, although occasionally they were simple.
- Venation in their leaves was open, and closure was rare.
- They had syndetocheilic-type stomata, where the two guard cells and subsidiary cells were derived from a single mother cell.
- The walls of epidermal cells had sinuous, curvy patterns.
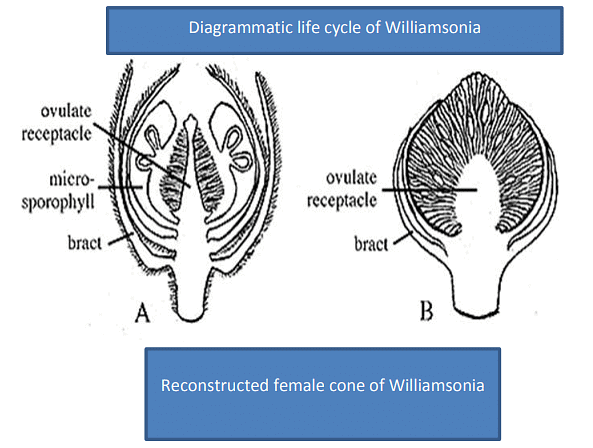
- Reproductive organs were organized as hermaphrodite "flowers" (e.g., Cycadeoidea) or unisexual "flowers" (e.g., Wielandiella), which were protected by multiple bracts.
- These "flowers" developed in the axils of leaves.
- Male reproductive organs were arranged in a whorl and could be free or fused, as well as entire or pinnately compound.
- Microsporangia were present abaxially in the form of synangia.
- Microsporophylls sometimes surrounded megasporophylls, forming hermaphrodite "flowers."
- Ovules were numerous, stalked, and borne on conical, cylindrical, or dome-shaped receptacles.
- Many inter-seminal bracts were present on the receptacle containing the ovules.
- Scales or bracts united at the end to form a shield through which the micropyle protruded.
- The seeds of Bennettitales were dicotyledonous.
These features highlight the unique characteristics of Bennettitales and their significance in the Earth's history during the Mesozoic era.
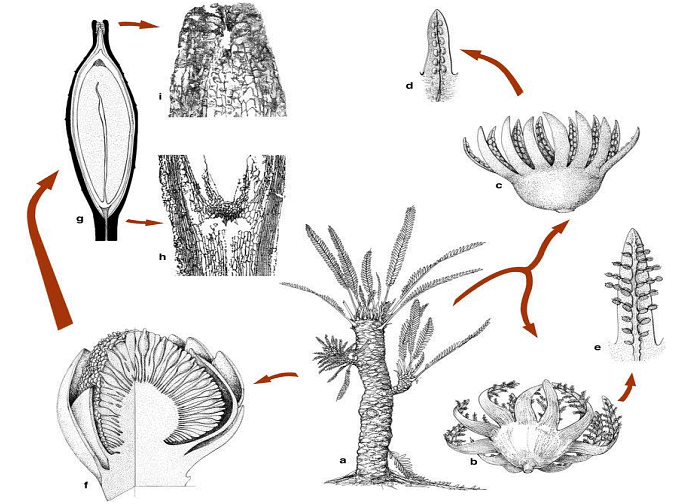
Resemblances of Bennettitales with Ferns:
- Multicellular Ramenta: Bennettitalean plants had multicellular ramenta covering their entire body, a feature also found in ferns.
- Direct Leaf Traces: Similar to ferns, Bennettitales had direct leaf traces.
- Scalariform Tracheids: The presence of scalariform tracheids, which are also seen in ferns, is another similarity.
- Large Pith: Bennettitales had a large pith, a characteristic shared with ferns.
Resemblances of Bennettitales and Cycads:
- Structure of Fronds: Bennettitales resemble cycads in the structure of their fronds.
- Short Stems with Persistent Leaf Bases: They share the presence of short stems covered with an armor of persistent leaf bases, which is a feature seen in cycads.
- Barrel-Shaped Trunk: Both groups have a barrel-shaped trunk.
- Cortex, Wood, and Pith in the Stem: They both possess a thick cortex, relatively thin wood, and a large pith in the stem.
- Manoxylic Wood: The type of wood, manoxylic wood, is common to both groups.
- Monocolpate Pollen Grains: Bennettitales have monocolpate pollen grains, a feature also observed in cycads. Monocolpate means having a single furrow-like aperture (colpus) in the pollen grain.
- Orthotropous Ovules: Both Bennettitales and cycads share orthotropous ovules, meaning the ovules are straight with the micropyle and the chalaza at opposite ends.
- Dicotyledonous Embryo: They both have dicotyledonous embryos.
While these resemblances suggest similarities between Bennettitales and both ferns and cycads, there are also fundamental differences between Bennettitales and cycads. These differences include aspects of their reproductive organs, such as the structure of "flowers," the position of ovules, and other characteristics that set them apart. Due to these dissimilarities, it is challenging to establish a direct phylogenetic connection between the two groups.
Andrews (1961) concluded that both Bennettitales and cycads likely evolved independently along separate evolutionary lines due to the significant differences between them, despite some shared features.
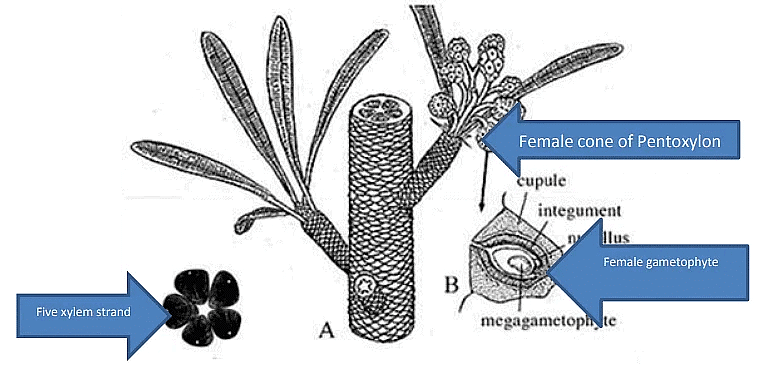
Pentoxylon
Habit : small trees or shrubs that possessed long and short shoots.Stem : called Pentoxylon because it frequently showed five segments of triangular vascular tissue arranged in a ring around a central ground tissue. The primary xylem is mesarch and secondary xylem is pycnoxylic ring around a central ground tissue. The primary xylem is mesarch and secondary xylem is pycnoxylic (resembling the wood of conifers). The short and long shoot are covered by an armour of spirally arranged leaf bases. The short shoots often terminated in a crown of spirally arranged Nipaniophyllum-type of leaves. Besides foliage leaves some short shoots were terminated by clusters of ovulate cones or pollen organs.
Discovery of Pentoxylales
The group known as "Pentoxyleae" was discovered and named by the renowned Indian Paleobotanist Professor Birbal Sahni in 1948. These fossil plants were found in the Rajmahal Hills in the Amrapara District of Eastern Bihar, India, indicating their existence during the Jurassic Period.
General Characteristics of Pentoxylon:
Stem (Pentoxylon):
- Exhibits short- and long-shoot morphology similar to Ginkgo.
- Diameter ranges from 3 mm to 2 cm.
- Each stele (vascular cylinder) has its own cambium.
- Cambium is uniformly active in young stems but becomes more active toward the center in mature stems, resulting in eccentric secondary wood growth.
- Primary xylem is mesarch.
- Secondary xylem is pycnoxylic, resembling coniferous wood, and is arranged in five or six wedges around the mesarch primary xylem strand.
- Short and long shoots are covered by spirally arranged leaf bases.
- Short shoots often terminate in a crown of spirally arranged Nipaniophyllum-type leaves.
- Some short shoots terminate in clusters of leaves.
- Presence of periderm and coniferous-type secondary xylem are indirect indications of its tree-like nature.
- The stem's height is hypothetically estimated to be 2-6 meters with robust and crinkled characteristics.
Leaves (Nipaniophyllum):
- Petiolate strap-shaped leaves with a prominent midrib.
- Closely resemble the form-genus Taeniopteris in general features.
- Structurally preserved leaves are referred to as Nipaniophyllum.
- Leaves are long, narrow (similar to Taeniopteris), and bear Syndetocheilic stomata.
- Prominent mid-rib and secondary veins arise at right angles and remain unbranched to the leaf margin.
- Vascular bundles are arranged in rows, each surrounded by a sclerenchymatous bundle sheath.
- Shows a diploxylic condition with central protoxylem, a larger centripetal primary metaxylem mass, and a small arc of centrifugal mass.
- The stiff, coriaceous leaves are borne in bunches in the apical portion of slender shoots.
- The plants were deciduous, dropping off leaves entirely.
Male Flower (Sahnia):
- Pollen organs were born in clusters on short shoots.
- Present terminally on the shoot and fused basally in a shallow disc.
- As many as 24 pollen-bearing organs have been reported.
- Consisted of a receptacle from which numerous microsporophylls or microsporangiophores arose.
- Stalks gave rise to secondary laterals that terminated in several stalked pollen sacs.
- Each microsporophyll possessed many pear-shaped, unilocular sporangia.
- The terminal position of the sporophyll was also occupied by a sporangium.
- Some details of the male-bearing structure are still unknown.
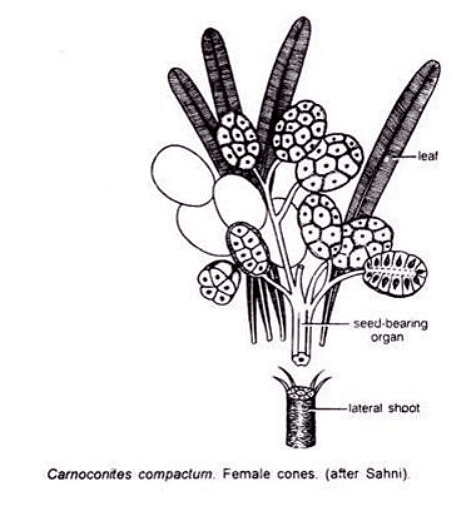
Female Cone (Carnoconites):
- Ovules aggregated into strobili.
- These strobili were mulberry-like and reached lengths of about 1.8 cm in Carnoconites compactum and 3 cm in C. laxum.
- They were narrower in C. laxum.
- Attached to short shoots.
- Unisexual (possibly too specialized to be an ancestor to angiosperms).
- About 20 sessile ovules were seen attached to the receptacle, with no inter-seminal scales.
- No sporophyll-like structures were reported.
- The ovules were covered by a single integument.
- The nucellus was free from the integument.
- Ovules were almost completely surrounded by a cupule.
- The integument had an inner sclerenchymatized layer and a fleshy outer layer.
Affinities of Pentoxylales
The affinities of Pentoxylales, an ancient group of gymnosperms, are discussed with various plant groups as follows:
Affinities with Cycadales:
- Pentoxylales share similarities with Cycadales, another group of gymnosperms.
- Both groups possess direct leaf traces in Pentoxylales and some cycad seedlings.
- They exhibit similarities in leaf trace anatomy, showing a diploxylic nature of their vascular bundles.
- Both groups have haplocheilic stomata in their leaves.
- Similarities in the nature of wood and pittings are observed between Pentoxylales and Cycadales.
Affinities with Conifers:
- Pentoxylales also display resemblances to conifers, another gymnosperm group.
- Shared characteristics include the presence of pycnoxylic wood.
- Both groups have tracheids with circular bordered pits arranged in an uniseriate or bi-senate manner.
- Uniseriate wood rays are common in both Pentoxylales and conifers.
Affinities with Medullosaceae:
- Pentoxylales share similarities with members of the Medullosaceae family within the Palaeozoic Pteridospermales (e.g., Medullosa).
- Both groups possess a polystelic primary vasculature in their stems.
- The secondary wood of Pentoxylon is pycnoxylic, a characteristic also found in some species of Medullosa.
- Similarities in their mode of branching and the nature of their steles are observed.
- Coniferous-type pittings are present in the stems of Pentoxylon and some Medullosa species.
Affinities with Bennettitales:
- Pentoxylales share resemblances with Bennettitales, another extinct group of gymnosperms.
- Both groups have syndetocheilic stomata in addition to haplocheilic ones.
- The diploxylic nature of vascular bundles is observed in both Pentoxylales and Bennettitales.
- Both groups exhibit whorled micro-sporangiophores.
- Some superficial resemblances exist between the male flowers of both groups.
- The manner in which ovules are borne in Pentoxylales is similar to that of Bennettitales, but inter-seminal scales found in Bennettitales are absent in Pentoxylales.
- Both groups share several common characteristics in their dwarf shoots.
- The presence of direct leaf traces also brings the two groups closer to each other.
These diverse affinities suggest that Pentoxylon is a unique gymnosperm group with features that combine elements from different gymnosperm lineages. Its secondary wood characteristics, leaf structures, reproductive organs, and mode of attachment of ovules make Pentoxylon a distinct and intriguing gymnosperm group with a synthetic nature.
|
179 videos|143 docs
|



ppt
,Previous Year Questions with Solutions
,Free
,study material
,Summary
,Type of Fossils and their Study Techniques | Botany Optional for UPSC
,Type of Fossils and their Study Techniques | Botany Optional for UPSC
,shortcuts and tricks
,Semester Notes
,video lectures
,Viva Questions
,practice quizzes
,Exam
,MCQs
,Extra Questions
,Important questions
,past year papers
,Type of Fossils and their Study Techniques | Botany Optional for UPSC
,Sample Paper
,mock tests for examination
,Objective type Questions
;


Type of Fossils and their Study Techniques Free PDF Download
Importance of Type of Fossils and their Study Techniques
Type of Fossils and their Study Techniques Notes
Type of Fossils and their Study Techniques UPSC Questions
Study Type of Fossils and their Study Techniques on the App
|
© EduRev
|
Education Revolution
|



|





