Incompatibility and Male Sterility and their Utilization in Crop Improvement - 1 | Agriculture Optional Notes for UPSC PDF Download
Self-Incompatibility
Self-incompatibility and sterility are two mechanisms that promote cross-pollination and are observed in over 300 species across 20 different angiosperm families.
Definition
In self-incompatible plants, the flowers produce functional pollen grains that are capable of fertilization. However, these pollen grains fail to fertilize either the same flower or any other flower on the same plant.
This failure can occur in several ways:
- Some self-incompatible pollen grains may not germinate on the stigmatic surface.
- Others may germinate but fail to penetrate the stigmatic surface.
- Some pollen grains may produce a pollen tube that enters through the stigmatic surface, but their growth is too slow. Consequently, by the time the pollen tube reaches the ovule, the flower has already wilted.
- In some cases, fertilization might occur, but the resulting embryo degenerates at an early stage.
Reason
Self-incompatibility is believed to be a result of biochemical reactions, although the exact nature of these reactions is not yet fully understood.
Classification of self incompatibility
According to Lewis (1954) the self incompatibility is classified as follows:
Heteromorphic System
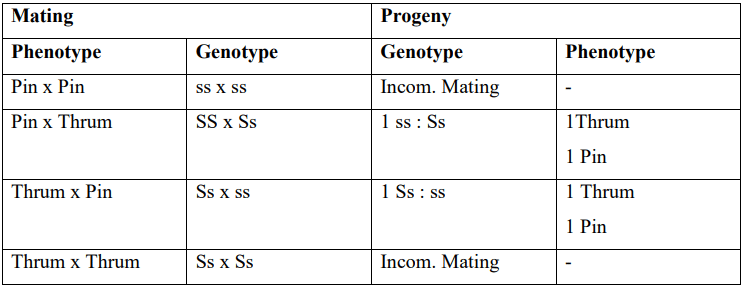
In the heteromorphic system, there are distinct differences in the appearance of the flowers. For instance, in the case of Primula sp, there are two flower types: PIN and THRUM. PIN flowers have a long style and short stamens, while THRUM flowers have a short style and long stamens. This contrast in flower morphology is referred to as Distyly. In Distyly, the only compatible mating is between PIN and THRUM flowers. The relative positioning of anthers is determined by a single gene, S/s, where the recessive allele "s" leads to PIN flowers, and heterozygotes "Ss" result in THRUM flowers.
Homozygous dominant individuals with "SS" do not exist as they are lethal. The incompatibility of pollen is influenced by the genotype of the plant producing it, with the allele "S" being dominant over "s". This system is also known as the heteromorphic-sporophytic system. Pollen grains from PIN flowers have a genotype of "ss" and display incompatibility reactions accordingly. On the other hand, THRUM flowers produce two types of gametes, "S" and "s", but all of them appear as "S" phenotypically. When PIN and THRUM flowers mate, they produce Ss and ss offspring in equal frequencies.
This system has limited significance in crop plants but can be found in sweet potatoes and some varieties of wheat.
TRISTYLY is known in some plants like Lythrum salicaria. In this case the style of the flower may be short, long or medium length.
Homomorphic System
In this case, the incompatibility is not linked to any visible differences in flower morphology. The reaction of pollen incompatibility can be determined either by the genotype of the plant on which the pollen is produced, known as sporophytic control, or by the genotype of the pollen itself, referred to as gametophytic control.
Gametophytic system
First identified by East and Mangelsdorf in 1925 in Nicotiana sanderae, this system is characterized by the fact that the pollen's incompatible reaction is dictated by its own genetic makeup and is not influenced by the plant on which the pollen is generated. Genetically, the incompatibility reaction is controlled by a single gene with multiple alleles. Examples of species that exhibit this system include Trifolium, Nicotiana, Lycopersicon, Solanum, and Petunia. Codominance is the assumed mode of inheritance in this system.
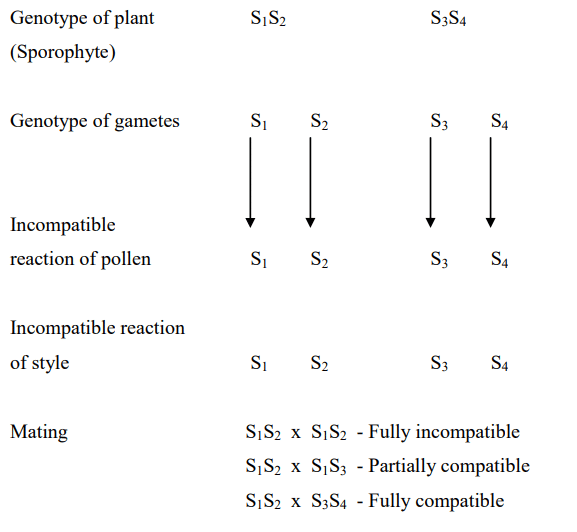
Sporophytic System
In this system, self-incompatibility is controlled by a single gene labeled as S, which possesses multiple alleles. Brassica oleracea, for example, has more than 30 known alleles for this gene. In this system, the dominance of alleles is assumed.
The self-incompatibility reaction is determined by the genetic makeup of the plant on which the pollen grain is produced and is not influenced by the genotype of the pollen itself. This system is notably complex, with alleles displaying dominance, co-dominance, or competition. The sporophytic system, as first reported by Hugues and Babcock in 1950 in Crepis foetida and by Gerstal in Parthenium argentatum, is found in various plants, including radish, brassicas, and spinach.
Lewis has summarized the characteristics of the sporophytic system as follows:
- Reciprocal differences are common.
- Incompatibility can occur with the female parent.
- A family can consist of three incompatibility groups.
- Homozygotes are a regular part of the system.
- An incompatibility group may contain two genotypes.
Mechanism of Self Incompatibility
This is quite complex and is poorly understood. The various phenomena observed in Self incompatibility is grouped into three categories.
- Pollen – Stigma interaction
- Pollen tube – Style interaction
- Pollen tube – Ovule interaction
Pollen – Stigma interaction
Incompatibility mechanisms become active shortly after pollen grains land on the stigma, typically inhibiting pollen germination. In the past, it was believed that the binucleate condition of pollen in gametophytic systems and the trinucleate condition in sporophytic systems were responsible for self-incompatibility. However, subsequent observations revealed that these conditions were not the primary cause of self-incompatibility.
Under the homomorphic system of incompatibility, there are variations in the stigmatic surface that hinder pollen germination. In gametophytic systems, the stigma surface is characterized by plumose structures, featuring elongated receptive cells, often referred to as a wet stigma. In this scenario, the pollen grain begins to germinate upon reaching the stigma, and the incompatibility reaction occurs at a later stage.
Conversely, in sporophytic systems, the stigma is papillate and dry, covered by a hydrated layer of protein known as a pellicle. This pellicle plays a crucial role in the incompatibility reaction. Within a few minutes of reaching the stigmatic surface, the pollen releases exine exudates, which are composed of either protein or glycerol protein. These exudates interact with the pellicle and trigger the formation of callose, a substance that further impedes the growth of the pollen tube.

Pollen Tube – Style interaction
In incompatible combinations, after pollen grains have germinated and the pollen tube has penetrated the stigmatic surface, the growth of the pollen tube is hindered within the style. This phenomenon can be observed in plants such as Petunia, Lycopersicon, and Lilium. In these cases, the synthesis of proteins and polysaccharides within the pollen tube ceases, eventually causing the pollen tube to burst and resulting in the death of its nuclei.
Pollen Tube – Ovule interaction
In Theobroma cacao pollen tube reaches the ovule and fertilization occurs but the embryo degenerates later due to some biochemical reaction.

|
4.61/5 Rating |

|
Dec 22, 2024 Last updated |

|
Explore Courses for UPSC exam
|
|
Semester Notes
,Sample Paper
,Important questions
,Viva Questions
,shortcuts and tricks
,video lectures
,Previous Year Questions with Solutions
,Incompatibility and Male Sterility and their Utilization in Crop Improvement - 1 | Agriculture Optional Notes for UPSC
,mock tests for examination
,ppt
,past year papers
,Summary
,Objective type Questions
,MCQs
,Free
,practice quizzes
,Exam
,Extra Questions
,Incompatibility and Male Sterility and their Utilization in Crop Improvement - 1 | Agriculture Optional Notes for UPSC
,study material
,Incompatibility and Male Sterility and their Utilization in Crop Improvement - 1 | Agriculture Optional Notes for UPSC
;






Incompatibility and Male Sterility and their Utilization in Crop Improvement - 1 Free PDF Download
Importance of Incompatibility and Male Sterility and their Utilization in Crop Improvement - 1
Incompatibility and Male Sterility and their Utilization in Crop Improvement - 1 Notes
Incompatibility and Male Sterility and their Utilization in Crop Improvement - 1 UPSC Questions
Study Incompatibility and Male Sterility and their Utilization in Crop Improvement - 1 on the App
|
© EduRev
|
Education Revolution
|



|




