Protochordata: Overview | Zoology Optional Notes for UPSC PDF Download
Understanding Chordata and Its Fundamental Characteristics
The classification of organisms into various phyla is a fundamental aspect of biology. Within the Animal Kingdom, one of the major phyla is Chordata, which was formally established by Balfour in 1880. The name "Chordata" is derived from the Greek words "chorde," meaning a string or cord, and "ata," meaning bearing. This phylum is characterized by a distinctive, supportive structure found along the back, known as the notochord. This shared characteristic sets chordates apart from the rest of the animal kingdom, often referred to as "non-chordates" or "invertebrates" due to their lack of a notochord or backbone.
Key Features of Chordates
Chordates share three fundamental morphological features at some point during their life cycle. These features are:
- Dorsal Hollow Nerve Cord: The central nervous system of chordates is positioned dorsally within the body. It takes the form of a longitudinal, hollow or tubular nerve cord situated just above the notochord and extending throughout the body. This nerve cord, or neural tube, develops from the dorsal ectodermal neural plate of the embryo and contains a cavity called the neurocoel. Notably, there are no distinct ganglionic enlargements along this nerve cord. The primary function of the nerve cord is to integrate and coordinate various bodily activities. In vertebrates, the anterior portion of the nerve cord undergoes specialization to form the cerebral vesicle or brain, which is protected by a bony or cartilaginous cranium. The posterior segment of the nerve cord develops into the spinal cord, safeguarded by the vertebral column.
- Notochord or Chorda Dorsalis: The notochord is an elongated, rod-like, and flexible structure that runs the length of the body. Positioned immediately beneath the nerve cord and just above the digestive canal, it originates from the endodermal roof of the embryonic archenteron. Structurally, the notochord is composed of large vacuolated cells. In adult vertebrates, it may be enclosed or replaced by the vertebral column.
- Pharyngeal Gill Slits: Chordates, at some point in their life cycle, develop a series of paired lateral gill clefts or gill slits that penetrate the pharyngeal wall behind the mouth. These structures are referred to by various names, such as pharyngeal, branchial, and visceral clefts or pouches. These gill slits serve primarily for the passage of water from the pharynx to the external environment, enabling respiration through gills. In addition, the water current assists in filter feeding by trapping food particles in the pharynx. While gill slits remain functional throughout life in protochordates and lower aquatic vertebrates, they are modified or lost in adult stages of higher vertebrates, which acquire pulmonary respiration.
Evolutionary Significance
Although the three common features—dorsal hollow nerve cord, notochord, and pharyngeal gill slits—appear during early embryonic development in all chordates, they often undergo modifications or are lost in the adult stage. In many vertebrates, the notochord disappears during development, while the nerve cord and pharyngeal clefts or their derivatives persist into adulthood. These shared characteristics are believed to have been present in ancestral chordates and set chordates apart from all other animals, pointing to their common ancestry.
Origin and Evolutionary History of Chordata
While the characteristics of modern chordates, including lower forms, are well-documented, the origin of this phylum remains shrouded in mystery. Determining the exact lower forms from which chordates evolved has proven challenging, primarily because their early ancestors were likely soft-bodied and have left no definitive fossil evidence. These ancestral forms are believed to predate the Cambrian period, as the oldest known vertebrate fossils have been discovered in late Cambrian strata. The prevalent scientific view is that chordates have evolved from invertebrates. Several theories have been proposed to explain the origin of chordates from non-chordate groups, but they have limitations and are not entirely satisfactory.
- One theory suggests that chordates descended from Echinodermata. This hypothesis is based on the striking similarities between the larvae of echinoderms (bipinnaria) and hemichordates (tornaria), providing strong evidence for a common ancestry. It is proposed that free-swimming auricularian larvae of some ancestral echinoderms evolved into chordates through a process called paedomorphosis. In paedomorphosis, the larval stage is prolonged, bypassing metamorphosis and sexual reproduction.
- Many zoologists, including Romer, Berril, and Barrington, now support the deuterostome lineage theory of chordate evolution. According to this theory, the phyla Echinodermata, Hemichordata, and Chordata share a common ancestry based on embryological and biochemical evidence. Protochordates serve as the connecting link between early chordate ancestors and vertebrates. The differentiation likely occurred long before the Cambrian period. The earliest traces of vertebrates have been found in late Cambrian and Ordovician rocks, with various fish species appearing during the Silurian period. The subsequent geological periods witnessed the evolution of amphibians, reptiles, birds, and mammals.
Major Subdivisions of Phylum Chordata
Phylum Chordata encompasses a diverse array of groups, each displaying varying degrees of relationships to one another. Taxonomically, these groups are traditionally organized into larger functional divisions or subdivisions based on specific structures or characteristics. These subdivisions may be classified under different ranks depending on the system of classification used. Here, we outline the major subdivisions of Phylum Chordata:
Subphyla and Classes: Phylum Chordata is initially divided into three or four primary subdivisions, known as subphyla, based on the presence or absence of a notochord. These include:
- Subphylum 1: Hemichordata (or Adelochordata)
- Subphylum 2: Urochordata (or Tunicata)
- Subphylum 3: Cephalochordata
- Subphylum 4: Vertebrata
Protochordata and Euchordata: The first two subphyla (Urochordata and Cephalochordata) are marine, relatively small, and lack a vertebral column. They are collectively known as non-vertebrate or invertebrate chordates or protochordates, as they are considered early, primitive chordates closely related to the ancestral chordate lineage.
Acrania and Craniata: Protochordate subphyla lack a head and a cranium and are referred to as Acrania. In contrast, the subphylum Vertebrata (subdivision Euchordata) possesses a distinct head and cranium and is known as Craniata.
Agnatha and Gnathostomata: Within the Vertebrata (Craniata), a widely accepted division separates them into two major sections: Agnatha and Gnathostomata. Agnatha includes species that lack true jaws and paired appendages. These agnathans encompass a small number of primitive but highly specialized fish-like forms, such as the extinct ostracoderms and modern cyclostomes.
Pisces and Tetrapoda: Among the Gnathostomata, a fundamental division recognizes two superclasses: Pisces and Tetrapoda. Pisces includes strictly aquatic forms with paired fins, primarily fish species. Sometimes, this group may also include fish-like agnathans. The superclass Tetrapoda is comprised of four-legged land vertebrates, including amphibians, reptiles, birds, and mammals.
Anamnia and Amniota: Another method of grouping vertebrates focuses on a significant advancement in embryological development—the presence of the amnion membrane, allowing the laying of eggs on land. Animals with this membrane belong to the group Amniota, which includes classes Reptilia, Aves, and Mammalia. Those lacking this membrane are categorized as Anamniota, encompassing cyclostomes, fishes, and amphibians. Anamniotes are sometimes referred to as lower vertebrates, while amniotes are considered higher vertebrates.
Phylum Hemichordata
The Phylum Hemichordata represents a group of organisms that are considered to be the lowest chordates, and they hold significant phylogenetic importance as they provide a link between chordates and non-chordates, particularly echinoderms.

Hemichordates have unique anatomical features, and their free-swimming larval stage, known as tornaria, bears a striking resemblance to the larval stages of certain echinoderms.
Classification of Phylum Hemichordata
- Historically, the study of Hemichordata has undergone several changes in classification. The earliest record of a hemichordate was made by Eschscholtz in 1825, and he named the animal Ptychodera, initially considering it a holothurian. Another member, Balanoglossus clavigerus, was added to the group by Delle Chiaje in 1829. Many hemichordates were subsequently described, and they were grouped under the name "Enteropneusti," as proposed by Gegenbaur in 1870. Bateson, in 1885, studied the embryology of some enteropneusts and introduced the name "Hemichordata" to replace "Enteropneusta."
- The tornaria larva, once believed to be an echinoderm larva, was described by Johannes Miller, Krohn, Agassiz, and others. Metchnikoff in 1870 regarded tornaria as the larval stage of Enteropneusta. Kowalevsky made significant contributions to the accurate description of Balanoglossus. The hemichordate with an elongated proboscis was named Saccoglossus by Schimkewitsch in 1892, although Spengel treated this animal as a subgenus, Dolichoglossus, under the genus Balanoglossus in 1893. Van der Horst in 1939 considered Saccoglossus and Dolichoglossus as synonyms.
- The classification of hemichordates was further complicated by the inclusion of colonial organisms like Rhabdopleura and Cephalodiscus, which were initially regarded as bryozoans but later suggested to belong to Hemichordata. Sars and Allman recorded Rhabdopleura as a member of Bryozoa, and Lankester introduced the term "Pterobranchia" for Rhabdopleura, although it was still considered a subdivision under Bryozoa. Another similar animal, Cephalodiscus, was recognized by Mcintosh in 1882 and also regarded as a bryozoan. Fowler removed Rhabdopleura from Bryozoa and suggested its inclusion under Hemichordata.
Important Features
- Hemichordates are bilaterally symmetrical deuterostome coelomates.
- They can be solitary (enteropneusts) or colonial (pterobranchs).
- The soft body is divided into three regions: proboscis (protosome), collar (mesosome), and trunk (metasome), reflecting a tri-coelomate organization internally.
- The preoral buccal diverticulum is considered the stomochord rather than the notochord.
- Their digestive tube is complete and can be straight or U-shaped.
- Pharyngeal slits are present or absent, and when present, they vary in number and are found in the trunk.
- Hemichordates have a heart and kidney located in the proboscis (preoral lobe) and are supported by the buccal diverticulum (stomochord).
- The nervous system is intra-epidermal and consists of dorsal and ventral nerve cords that extend the length of the animal. This dorsal nerve cord in the collar region represents a true chordate feature.
- They exhibit separate sexes (gonochoristic), although some species are hermaphroditic.
- Gonads are located externally.
- Some hemichordates can undergo asexual budding, and buds form in the posterior end of the body.
- Fertilization can be external (in enteropneusts) or internal (in pterobranchs).
- Cleavage patterns can be holoblastic and radial (in enteropneusts) or radial and bilateral (in pterobranchs).
- Hemichordates may have a planktonic, ciliated tornaria larva that is non-feeding and utilizes stored reserves from the egg during development.
Fossil Record:
The fossil record of hemichordates is limited, with few data available except for Eocephalodiscus, which has been recovered from Upper Cretaceous strata.
Geographical Distribution:
Hemichordates are found in various seas, with a preference for warm and temperate waters. Most species inhabit the intertidal zone and shallow waters, while some are found at greater depths.
Habitat:
Hemichordates live in U-shaped burrows within sand, mud, rocks, and among seaweed.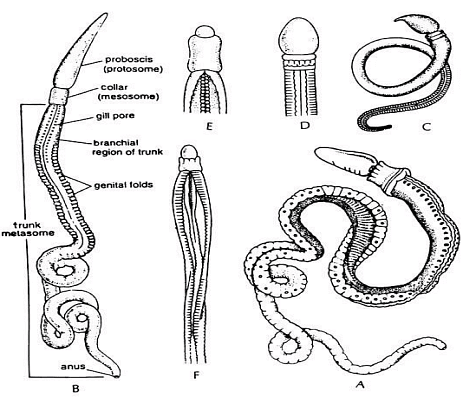 The Phylum Hemichordata includes two classes: Enteropneusta and Pterobranchia. Some authors have also included Planctosphaeroidea as a third class in their classifications.
The Phylum Hemichordata includes two classes: Enteropneusta and Pterobranchia. Some authors have also included Planctosphaeroidea as a third class in their classifications.
|
181 videos|346 docs
|

|
4.93/5 Rating |

|
Dec 22, 2024 Last updated |

|
Explore Courses for UPSC exam
|
|
Extra Questions
,Viva Questions
,practice quizzes
,Protochordata: Overview | Zoology Optional Notes for UPSC
,video lectures
,Important questions
,Free
,ppt
,MCQs
,mock tests for examination
,study material
,Exam
,Protochordata: Overview | Zoology Optional Notes for UPSC
,Protochordata: Overview | Zoology Optional Notes for UPSC
,Objective type Questions
,Summary
,past year papers
,Semester Notes
,Sample Paper
,shortcuts and tricks
,Previous Year Questions with Solutions
;






Protochordata: Overview Free PDF Download
Importance of Protochordata: Overview
Protochordata: Overview Notes
Protochordata: Overview UPSC Questions
Study Protochordata: Overview on the App
|
© EduRev
|
Education Revolution
|



|




