Spermatogenesis | Zoology Optional Notes for UPSC PDF Download

Introduction
Mammalian spermatogenesis is a meticulously coordinated and intricate cellular differentiation process, where a spermatogonial stem cell undergoes gradual transformation into a highly specialized haploid cell known as a spermatozoon. This progression involves three distinct categories of germinal cells—spermatogonia, spermatocytes, and spermatids—typically organized in concentric layers within the seminiferous tubules.
In adult mammals, spermatogenesis is an ongoing process marked by two distinct phases, each exhibiting specific morphological and biochemical alterations in the nuclear and cytoplasmic components.
The two phases include:
(i) formation of spermatids (mitosis and meiosis) and
(ii) spermiogenesis.
Formation of spermatids
This phase of spermatogenesis is further subdivided into three phases.
Multiplication phase
This stage is alternatively termed as the proliferation and renewal period for spermatogonia. In this phase, the diploid spermatogonia located at the outer edge of the seminiferous tubule undergo mitotic division, leading to the formation of spermatocytes. Additionally, these spermatogonia generate new stem cells and transition into a growth phase.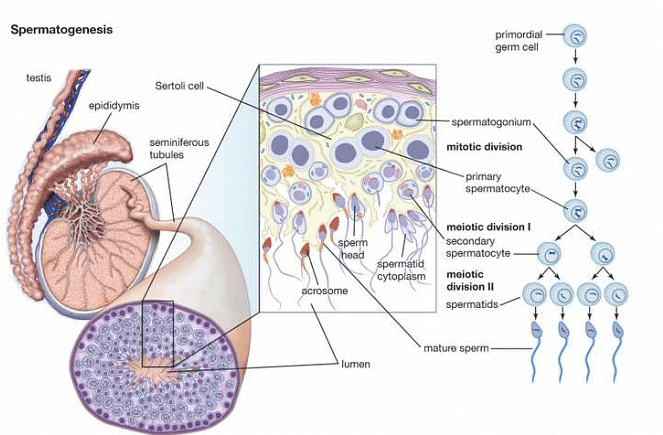
Growth phase
In this stage, there is a constrained expansion of spermatogonia, causing their size to double, and they are subsequently referred to as primary spermatocytes, maintaining their diploid status. Subsequently, these primary spermatocytes progress to the next stage, known as the maturation phase.
Maturation phase
The primary spermatocytes initiate the prophase of meiotic or maturation division during this phase. Meiotic prophase is a highly intricate process characterized by a systematic sequence of chromosomal rearrangements accompanied by molecular changes. In the course of meiosis, the nuclear DNA first duplicates, and each homologous chromosome undergoes pairing (synapsis) and longitudinal splitting into two chromatids, which remain connected by a common centromere.
Chiasma formation, involving the mutual exchange of some chromosome material between two non-sister chromatids of each homologous pair (tetrad) through crossing over, results in an almost limitless variety of combinations of paternal and maternal genes in any gamete. Ultimately, two chromosomes from each homologous pair (tetrad) migrate to opposite poles of the primary spermatocyte. At this point, each pole of the primary spermatocyte possesses a haploid set of chromosomes, enclosed by the nuclear membrane developed from the endoplasmic reticulum.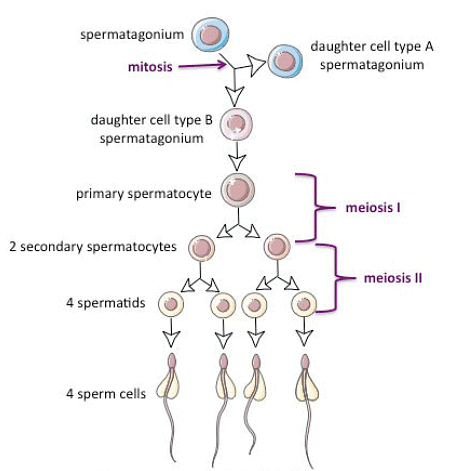
Typically, the first meiotic division is followed by the division of cytoplasm (cytokinesis), resulting in the division of each primary spermatocyte into two haploid secondary spermatocytes. Each secondary spermatocyte undergoes the second meiotic or maturation division, which is akin to a simple mitosis and produces four haploid spermatids. These spermatids are initially non-functional male gametes and need to undergo a multifaceted process of cytological and chemical transformations, commonly known as spermiogenesis, to become functional spermatozoa.
Spermiogenesis
The progression of spermatids undergoing alterations to give rise to spermatozoa is known as spermiogenesis. As a spermatozoon is an exceptionally active and mobile cell, achieving genuine mobility necessitates the elimination of unnecessary materials from developing spermatozoa, leading to a high degree of specialization in the sperm cell through a series of steps.
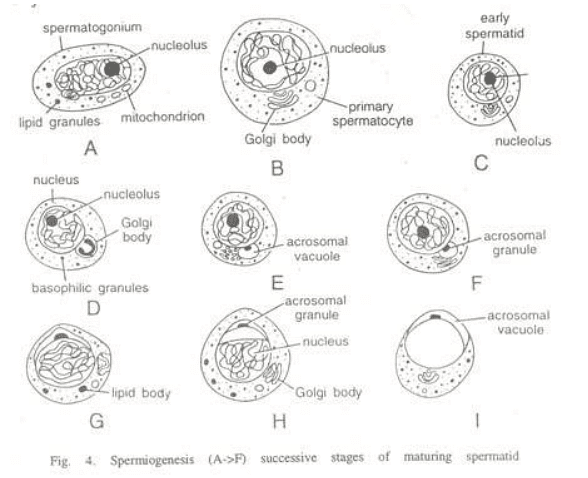
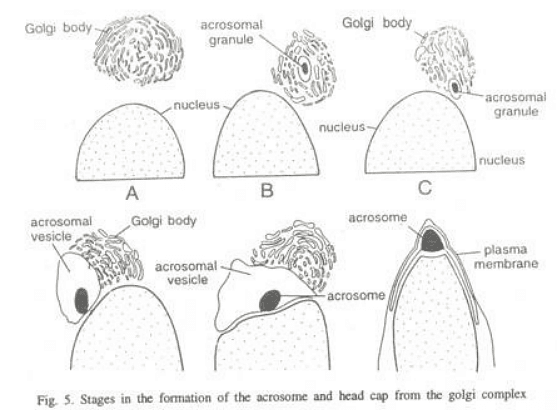
Spermiogenesis involves the formation of two primary components of the sperm, namely the head and the tail.
Formation of head
The transformation of the two primary components of the sperm head, namely the nucleus and acrosome, involves the following changes during spermiogenesis.
- Changes in the nucleus: Throughout spermiogenesis, the nucleus of the spermatid undergoes a reduction in size as it loses a significant amount of water from the nuclear cap. The chromosomes become densely packed into a smaller volume, and the entire ribonucleic acid is eliminated, leaving only the genetic material, the deoxyribonucleoprotein. This process involves the removal of material not directly involved in the transmission of hereditary characteristics. The spherical shape of the nucleus also elongates and narrows, adapting for propulsion in fluid mediums and penetration of the ovum, with the specific shape varying among different animals.
- Golgi phase: In the initial stages, the young spermatid has a round shape with a spherical nucleus. The Golgi apparatus secretes glycoprotein-rich granules, known as proacrosomic granules, which fuse to form a single large acrosomal granule attached to the nuclear membrane.
- Cap phase: The acrosomal granule flattens onto the nucleus of the spermatid, forming the head cap. The Golgi apparatus, responsible for secreting the acrosome, separates from the head cap and moves towards the opposite pole. Meanwhile, the centrioles, located near the nucleus on the side opposite the acrosomic cap, develop a flagellum.
- Acrosome phase: The definitive morphological features of the acrosome become clearly defined. The remaining part of the Golgi apparatus is gradually reduced and ultimately discarded from the sperm as "Golgi-rest," along with some cytoplasm. In the spermatozoon, the axial body or acrosomal core appears between the acrosomal granule and nucleus, extending itself from behind into the acrosomal granule. This body develops as the sperm approaches the egg and contains enzymes used to dissolve the egg membranes during the fertilization process.
Formation of the tail of the spermatozoon
Following the second meiotic division, the centrosome of a spermatid consists of two centrioles, structured as two cylindrical bodies lying perpendicular to each other. In the early stages of sperm metamorphosis, these centrioles relocate to a position just behind the future neck region of the sperm nucleus. A depression forms in the posterior surface of the nucleus, and one of the centrioles, known as the proximal centriole, is positioned in the depression with its axis approximately at right angles to the main axis of the spermatozoon. The other centriole, the distal centriole, takes a position behind the proximal one, aligning its axis with the longitudinal axis of the spermatozoon.
The distal centriole gives rise to the axis filament of the spermatozoon, serving as the basal granule. Most of the mitochondria of spermatids concentrate around the distal centriole and the upper part of the axial filament, forming the neck and middle piece of the spermatozoon's tail. In the middle piece of the sperm, the mitochondria lose their individuality through fusion to varying degrees. In mammals, the mitochondria join into one continuous body, spirally twisted around the proximal part of the axial filament and the proximal centriole.
In contrast, some animals lack a spiral arrangement of mitochondria, and instead, these mitochondria fuse together to create dense clumps known as mitochondrial bodies. The cytoplasm forms a condensed layer called the manchette around the periphery of the middle piece. The manchette also surrounds the posterior part of the head of the spermatozoa, except where it is not covered by the cap.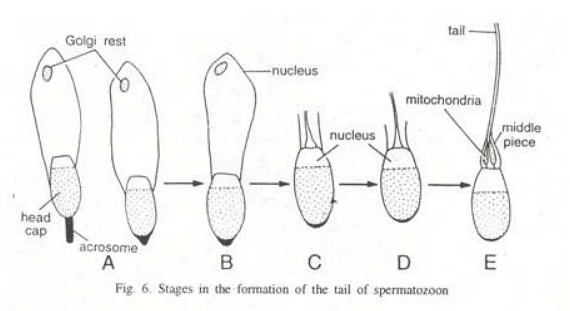
A dark ring known as the 'Ring Centriole' is observed at times at the posterior end of the middle piece of the sperm. The function of this ring centriole is not well understood, and it serves as the boundary between the middle piece and the principal piece of the sperm tail. In most animals, excluding mammals, the principal piece and tail piece of the sperm consist solely of the axial filament. However, in mammals, the axial filament of the principal piece is accompanied on its outer side by much thicker, wedge-shaped fibers that originate in the middle piece but do not extend to the end piece of the spermatozoon tail.
In mammalian sperm tails, these fibers of the axial filament are also surrounded by flattened bands, forming semi-circular ribs that articulate with each other on opposite sides of the sperm tail. The end piece of the tail contains only the axial filament, which remains covered with cytoplasm and the plasma membrane.
Biochemical Changes in Spermatogensis
Various biochemical processes take place during spermatogenesis, as outlined by Monesi in 1970:
- RNA synthesized during meiosis is expelled from the nucleus in both meiotic divisions and persists in the cytoplasm. Fully developed spermatozoa show no detectable amounts of RNA, with meiotic RNA likely being involved in the synthesis of acrosomal proteins and the flagellum.
- The synthesis of nuclear proteins is halted midway through spermiogenesis.
- All non-histone proteins in the nucleus are removed during spermiogenesis.
- Synthetic events occurring in spermiogenesis are likely regulated by stable RNA produced during the meiotic stages.
- The suppression of genetic activity in spermatids and spermatozoa might be influenced by a regulatory mechanism leading to the disappearance of activating proteins and RNA from the chromosomes.
- Histone molecules associated with DNA may have a protective function, stabilizing DNA against changes during their transit through the male and female reproductive tracts, as suggested by Taiwan et al. in 1989.
Control of Spermatogenesis
Spermatogenesis can be influenced by environmental or physiological factors, with temperature, light exposure, hormones, and psychological state playing significant roles depending on the organism. The pituitary gland is a key regulator of spermatogenesis, releasing specific gonadotrophin hormones. However, the activities of the pituitary gland and gonads in birds, rodents, and various other vertebrates are susceptible to environmental factors such as temperature, light conditions, and day length.
|
198 videos|351 docs
|
FAQs on Spermatogenesis - Zoology Optional Notes for UPSC
| 1. What is spermatogenesis? |  |
| 2. How are spermatids formed during spermatogenesis? | |
| 3. What happens during spermiogenesis? | |
| 4. What are the main stages of spermatogenesis? | |
| 5. How long does spermatogenesis take? | |


Sample Paper
,MCQs
,Previous Year Questions with Solutions
,shortcuts and tricks
,Extra Questions
,Important questions
,Free
,Spermatogenesis | Zoology Optional Notes for UPSC
,practice quizzes
,Spermatogenesis | Zoology Optional Notes for UPSC
,study material
,mock tests for examination
,Semester Notes
,ppt
,Exam
,Spermatogenesis | Zoology Optional Notes for UPSC
,past year papers
,Objective type Questions
,Summary
,video lectures
,Viva Questions
;


Spermatogenesis Free PDF Download
Importance of Spermatogenesis
Spermatogenesis Notes
Spermatogenesis UPSC Questions
Study Spermatogenesis on the App
|
© EduRev
|
Education Revolution
|



|





