Structure and Reproduction Fungi & Lichens | Botany Optional for UPSC PDF Download
| Table of contents |
|
| Thallus Organization and Reproductive Pattern in Fungi |
|
| Vegetative Thallus Organisation |
|
| Reproduction in Fungi |
|
| Structure and Reproduction in Lichen |
|
| Reproduction |
|
Thallus Organization and Reproductive Pattern in Fungi
- A thallus refers to the undifferentiated vegetative tissue found in various organisms across different groups, including algae, fungi, certain liverworts, lichens, and the Myxogastria. Many of these organisms were once collectively referred to as thallophytes, even though they are not closely related. A structure or organism resembling a thallus can be called thalloid, thallodal, thalliform, thalline, or thallose.
- In the context of a thallus, it typically denotes the entire body of a multicellular, non-moving organism that lacks the organization of tissues into distinct organs like leaves, roots, and stems, as seen in vascular plants. However, thalli may have structures that serve analogous functions to the vascular plant's organs, despite having different microscopic structures. For instance, thalli lack vascular tissue. In exceptional cases, like the Lemnoideae, which exhibit a structure resembling a vascular plant, it is described as having a thalloid structure.
- While a thallus is generally undifferentiated in terms of its internal anatomy, there can be visible and functional differences. For example, in kelp, the thallus can be divided into distinct regions, including the holdfast (which anchors the organism), the stipe (providing support to the blades), and the blades (responsible for photosynthesis).
- In the case of fungi, the thallus is commonly referred to as a mycelium. The term thallus is also frequently used to describe the vegetative body of a lichen, while in seaweed, it may be called a 'frond.' In certain non-thallophyte plants like clubmosses, horsetails, and ferns, the gametophyte is termed a 'prothallus.'
- Fungi enter a reproductive phase after a period of intensive growth, during which they produce and release large quantities of spores. Spores are typically single cells generated either through the fragmentation of the mycelium or within specialized structures such as sporangia, gametangia, sporophores, and more. Spores can be produced directly through asexual methods or indirectly through sexual reproduction. In fungal sexual reproduction, like other organisms, it involves the fusion of two nuclei when two sex cells (gametes) unite. Asexual reproduction is a simpler and more direct method.
Vegetative Thallus Organisation
The vegetative structures in most fungi, except for unicellular ones, consist of a network of fine filaments known as mycelium. Each individual filament is referred to as a hypha (plural: hyphae). In many fungi, the thallus (the fungal body) is divided into a vegetative region responsible for nutrient absorption and a reproductive region. Such thalli are called eucarpic. In contrast, some fungi lack this differentiation, and after a phase of vegetative growth, the entire thallus transforms into reproductive structures. These thalli are termed holocarpic, as seen in organisms like Synchytrium and Olpidium.
The mycelium represents the vegetative phase of a fungus and originates from a spore. Hyphae can be either branched or unbranched and exhibit a wide range of sizes, varying from a few microns to several meters in length. Hyphal thickness can range from 0.5 µ to 100 µ. Typically, hyphae are colorless, but in some species, they may have various colors due to differences in reserve food materials or pigments.
Each hypha is composed of an outer thin wall and a cavity filled with or lined by protoplasm. Different groups of fungi exhibit variations in hyphal structure. Oomycetes and Zygomycetes generally have non-septate hyphae (lacking crosswalls or septa), whereas Ascomycetes, Basidiomycetes, and Deuteromycetes have septate hyphae (with crosswalls). In aseptate hyphae, numerous nuclei are dispersed throughout the cytoplasm, and this type of vegetative structure is termed coenocytic.
In septate fungi (Ascomycetes, Basidiomycetes), septa have a single central pore (or occasionally more) that allows the passage of cytoplasm, organelles, and even nuclei from one cell to another. Despite the presence of septa, these fungi can be considered coenocytic organisms. Ascomycetes have a simple pore in the middle of the crosswall, while Basidiomycetes have a more complex structure known as the dolipore.

In aseptate hyphae, septa form when reproductive structures are delimited or when a wall is developed to seal off damaged areas of the hypha. Within septate or cellular hyphae, individual hyphal cells may contain one nucleus (uninucleate), two nuclei (binucleate), or many nuclei (multinucleate). A hyphal cell with genetically identical haploid nuclei is termed monokaryotic, whereas cells containing two genetically different but compatible haploid nuclei are called dikaryotic.
Basidiomycetes exhibit two phases in their life cycle: a monokaryotic phase in which hyphal cells are uninucleate, and a dikaryotic phase in which cells are binucleate. The corresponding mycelium stages are referred to as monokaryotic mycelium and dikaryotic mycelium, which are also known as primary and secondary mycelium, respectively.
The vegetative cells of most fungi possess a well-defined cell wall made primarily of chitin, while some may also contain cellulose. Slime molds are an exception as they lack a cell wall. The hyphal wall is composed of layers of microfibrils of chitin and cellulose distributed within an amorphous matrix. In the majority of fungi, chitin is the predominant component of the cell wall. In certain fungi, like oomycetes, cellulose makes up the majority of the cell wall, while in others, the cell wall consists of both chitin and cellulose. Additional substances, such as glucan, have also been identified in fungal cell walls.
Slime molds have a vegetative body composed of a slimy mass of naked protoplasm without an external wall.
Within the cell wall lies the protoplast, which is surrounded by the plasmalemma (cytoplasmic membrane) containing ergosterol (not cholesterol or phytosterol as in animals or plants). Cellular inclusions in fungi include endoplasmic reticulum, ribosomes, mitochondria, vacuoles, and nuclei. Fungal endoplasmic reticulum consists of two parallel unit membranes, while ribosomes are dispersed throughout the cytoplasm. Mitochondria in fungi resemble those in other organisms. Vacuoles in fungal cells are large and evident.
Fungal cells possess lomasomes, membrane-bound tubules or vesicles found between the plasmalemma and the cell wall, although their function remains unknown. Dictyosomes, which are typically rare, have been reported in oomycetes.
All fungi are eukaryotic, meaning they possess a nucleus enclosed by a double membrane. Fungal nuclei are small, typically ranging from 2–3 µm in diameter, while those in plants and animals are larger (at least 5–8 µm). Fungal nuclei are separated by a porous double membrane. During mitotic nuclear division, the nuclear envelope may constrict in the middle to separate two sister nuclei, a process known as karyochorisis. Chromatin in fungi is generally uniform, with limited differentiation into euchromatin and heterochromatin. In some fungi like yeast and Neurospora, chromosomes do not condense and become visible during mitosis.
Mitosis in fungi can vary widely depending on the species. In some fungi, including slime molds and non-filamentous fungi, the spindle forms inside the nucleus, which remains distinct, as the nuclear envelope does not break down as in plants and animals. Instead, it constricts to separate the two daughter nuclei.
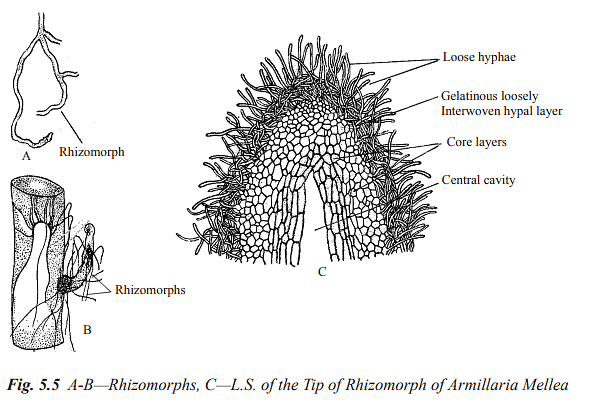
Fungal mycelium is typically a tangled mass of hyphae growing on or within a substrate. In certain fungi, such as Ascomycetes and Basidiomycetes, hyphae can organize into loosely or compactly woven structures known as hyphal strands. Two types of hyphal strands exist: prosenchyma (loosely woven) and pseudoparenchyma (closely packed with short cells). Hyphal cells from different strands may fuse through a process called anastomosis, forming a three-dimensional network of hyphae. Anastomosis facilitates the development of specialized structures like sclerotia, fruiting structures (stromata or sporocarps), and rhizomorphs.
Sclerotia are tough, often rounded resting bodies composed of pseudoparenchyma and are capable of surviving adverse conditions. Stromata are complex, organized structures found in ascomycetes and basidiomycetes fungi, where spores are produced sexually. Rhizomorphs are elongated, root-like strands that facilitate perennation and vegetative propagation, primarily in Basidiomycetes.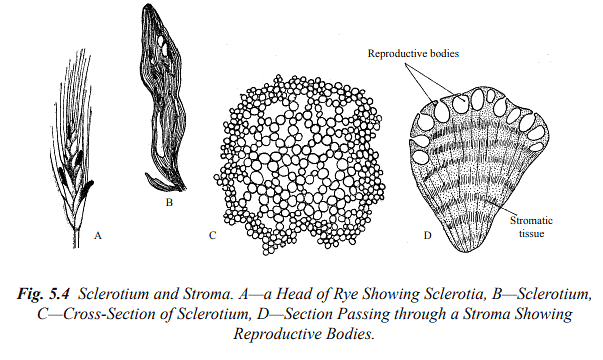
It's important to note that fungal cells and structures exhibit considerable diversity among different species and groups within the fungal kingdom.
Reproduction in Fungi
The reproduction in fungi takes place by following three methods:
- Vegetative Reproduction
- Asexual Reproduction
- Sexual Reproduction
Vegetative Reproduction
Fungal reproduction occurs through several methods:
(a) Fragmentation: In numerous fungi, the vegetative body breaks into one or more fragments, each of which can develop into a new individual when favorable conditions are present.
(b) Oidia: Some fungi produce small, rounded, or oval segments known as oidia when their hyphae break apart. Each oidium acts as an accessory spore and can germinate to form a new individual under suitable conditions.
(c) Chlamydospores: In certain fungi, specific cells within the hyphae develop thick walls and become chlamydospores. These are resting spores that can remain viable for extended periods. Chlamydospores are formed singly or in chains, either at the tips or along the length of the hyphae. When favorable conditions return, each chlamydospore can germinate to establish a new mycelium. Examples of fungi employing this method include Pythium, Mucor, and Fusarium.
(d) Fission: Unicellular fungi like yeasts reproduce vegetatively by splitting the vegetative cell into two equal daughter cells, followed by a constriction. Each daughter cell then matures into a new vegetative cell.
(e) Budding: In some fungi, such as yeasts, vegetative reproduction occurs through budding. The vegetative cell produces a small outgrowth called a bud, which eventually separates from the mother cell through constriction, forming a new cell. In some yeasts, multiple buds may form simultaneously and remain together, creating a chain of buds. This chain resembles mycelium and is referred to as pseudomycelium.
(f) Sclerotia: Many fungi produce sclerotia, which are durable, resistant structures. Each sclerotium is typically round and consists of outer thick-walled cells forming the rind, as well as inner parenchyma cells. Sclerotia can endure unfavorable conditions and, once favorable conditions return, they can germinate to produce vegetative hyphae, initiating new fungal growth.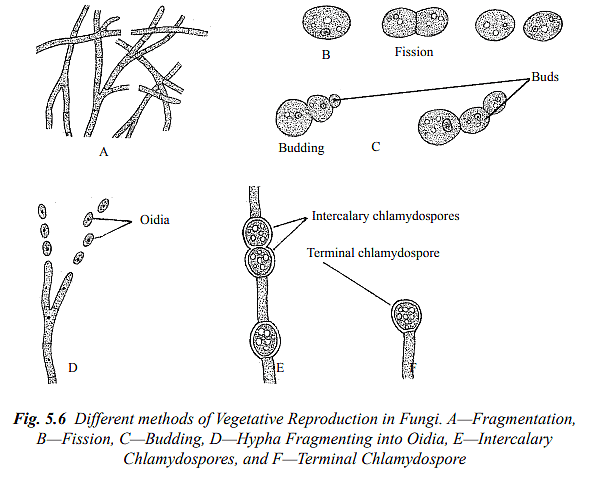
Asexual Reproduction
Asexual reproduction in fungi primarily occurs through the formation of spores, which are the most common types of reproductive units. These spores are typically small, usually unicellular, and are generated from the parent fungus without involving sexual processes. In some fungi, like Alternaria, the spores can be multicellular. Asexual spores, also known as mitospores, can be categorized into two main types based on their formation: sporangiospores and conidia.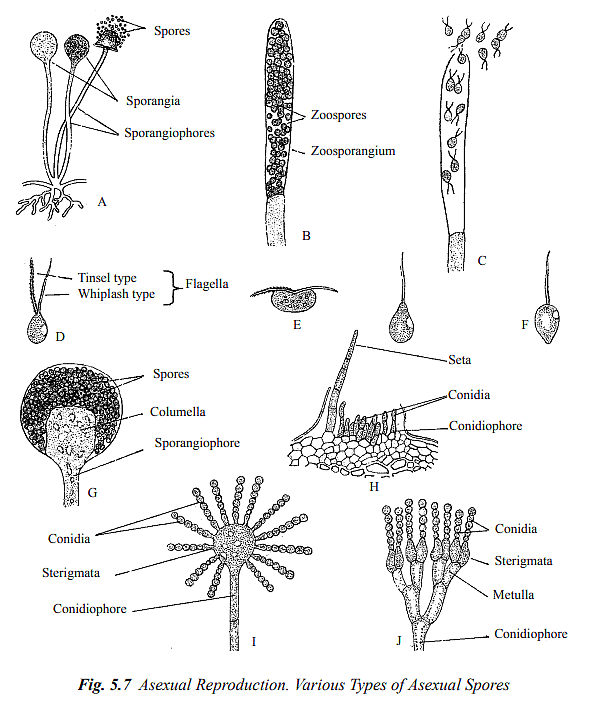
Sporangiospores: These spores are produced internally within specialized sac-like structures called sporangia. Sporangia are typically found on specialized hyphae known as sporangiophores. Depending on their motility, sporangiospores can be further divided into two subtypes:
Zoospores: These are motile sporangiospores produced in zoosporangia. They can move using flagella and are found in fungi like Saprolegnia, Pythium, and Phytophthora (Refer to Figure 5.7 C). Zoospores can possess one or two flagella with distinct characteristics (Refer to Figure 5.7 D).
Aplanospores: Aplanospores are non-motile sporangiospores formed endogenously. Fungi such as Mucor and Rhizopus produce aplanospores (Refer to Figure 5.7 A and G). These are unicellular spores dispersed by wind currents.
Conidia: In fungi belonging to the groups Ascomycetes, Basidiomycetes, and Deuteromycetes, asexual reproduction involves the production of non-motile spores called conidia. Conidia are typically formed externally at the tips of regular vegetative hyphae or specialized hyphae called conidiophores (Refer to Figure 5.7 I and J). These conidia can be present individually (e.g., Pythium, Phytophthora) or in chains (e.g., Aspergillus, Penicillium). Conidiophores may either be separate from each other or grouped together to create complex structures like ascervuli, pycnidia, synnemata, and sporodochia.
Sexual Reproduction
Asexual Reproduction: While many fungi primarily reproduce sexually, the members of mitosporic fungi, also known as Deuteromycetes or fungi imperfecti, lack known or observed sexual stages. In sexual reproduction in fungi, two nuclei of different parentage fuse together, and this fusion can occur in various forms, including motile or non-motile gametes, gametangia, or somatic cells of the thallus.
Homothallic and Heterothallic: Fungi can be homothallic, meaning they produce both male and female gametangia on the same mycelium and are capable of self-fertilization. In homothallic fungi, gametangia may either be clearly distinct as male and female structures (antheridia and oogonia) or morphologically similar and sexually undifferentiated.
In contrast, heterothallic fungi separate the sexes into different individuals:
Three Phases of Sexual Reproduction: Sexual reproduction in fungi occurs in three distinct phases that happen sequentially: plasmogamy, karyogamy, and meiosis.
- Plasmogamy: This is the initial phase where two protoplasts unite, bringing together two haploid nuclei of opposite sexes within a single cell.
- Karyogamy: In the second phase, two nuclei fuse, forming a diploid nucleus known as the synkaryon. In lower fungi, karyogamy follows plasmogamy almost immediately, while in higher fungi like Ascomycetes and Basidiomycetes, karyogamy is somewhat delayed, resulting in the formation of a dikaryon. As hyphae grow, these paired nuclei divide, eventually generating numerous dikaryotic cells.
- Meiosis: Meiosis, the third phase of sexual reproduction, promptly follows karyogamy. During meiosis, the diploid nucleus undergoes reduction division, reducing the chromosome number back to the haploid state. In certain fungi, such as oomycetes, the vegetative thallus is diploid, and meiosis occurs in the gametangia.
Sex Cells and Gametangia: The two merging protoplasts are known as sex cells or gametes, and they are produced within specialized structures in the thallus called gametangia.
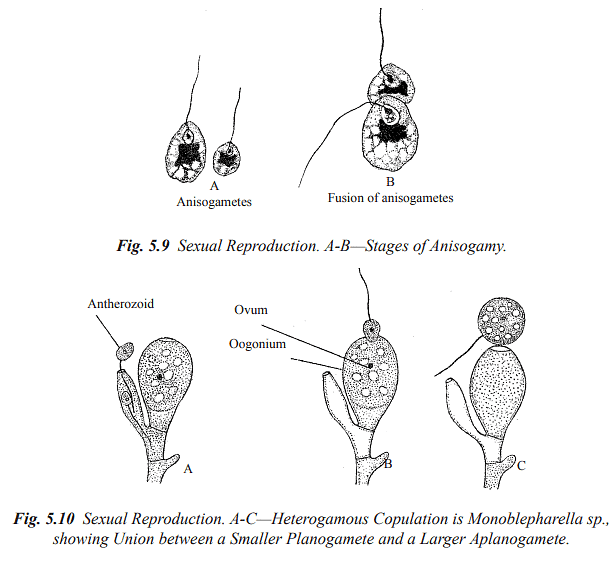
Isogamy: When the fusing gametes are morphologically identical, they are termed isogametes, and the fertilization process is referred to as isogamous. Isogametes can be flagellate (planogametes) or non-flagellate (aplanogametes).
Heterogamy: Heterogamous fungi produce different-sized and structured gametes. There are two subtypes:
- Anisogamy: This involves the fusion of two morphologically similar gametes that differ in size.
- Oogamy: Oogamy occurs when gametes are produced within morphologically distinct gametangia. The male gametangium, called the antheridium, is smaller, while the female gametangium, known as the oogonium or ascogonium, is larger.
Methods of Plasmogamy: The fusion of compatible nuclei through plasmogamy can happen through several methods:
Gametangial Contact: In this method, male and female gametangia come into direct contact with each other, allowing the male nucleus (or nuclei) to be transferred into the female gametangium through a pore or fertilization tubes.
Gametangial Copulation: Gametangial copulation involves the fusion of the entire contents of two gametangia through a pore or by dissolving contact walls, resulting in the formation of a common cell.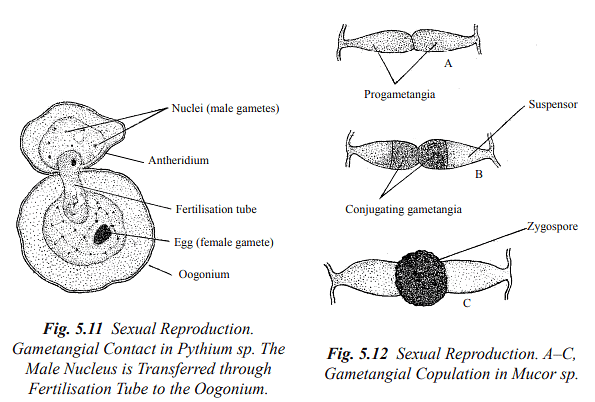
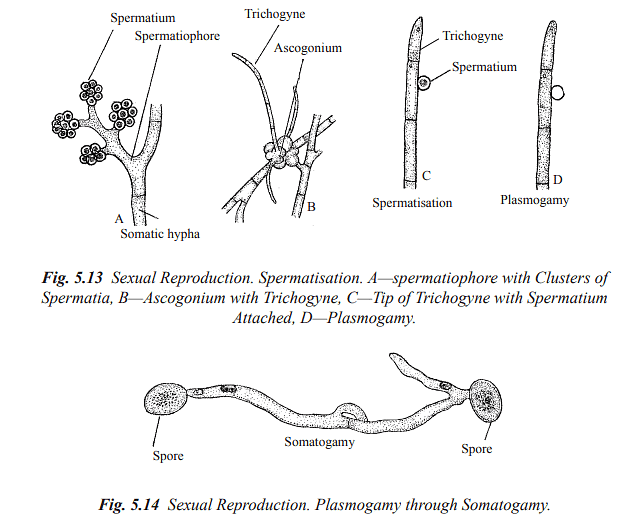
Spermatisation: Some fungi produce non-motile male cells called spermatia, which are carried by various means (e.g., wind, water, insects) to female gametangia, receptive hyphae, or somatic hyphae. These spermatia then transfer their contents into the receptive structure, forming a dikaryotic cell.
Somatogamy: In higher fungi where sex organs are absent, sexual reproduction occurs through somatogamy. This involves the anastomosis (fusion) of somatic hyphae that contain nuclei of different parentage.
Structure and Reproduction in Lichen
Structure of Lichens
Lichens can be categorized into three primary morphological groups based on their overall appearance: crustose, foliose, and fruticose lichen thalli.
- Crustose Lichens: Crustose lichens are firmly attached to their substrate to the extent that they are often inseparable from it. Common examples of crustose lichens include Graphis, Lecanora, Lecidia, and Haematomma.
- Foliose Lichens: Foliose lichens have a leaf-like thallus structure and are partially connected to their substrate through hair-like structures called rhizinae (see Figure 7.2 C). Physcia, Parmelia, Peltigera, and Collema are among the typical representatives of foliose lichens.
- Fruticose Lichens: Fruticose lichens are upright, shrub-like, pendent, and exhibit hair-like or strap-shaped structures attached only at their base (see Figure 7.2 D and E). Examples of fruticose lichens include Usnea, Cladonia, and Ramalina.
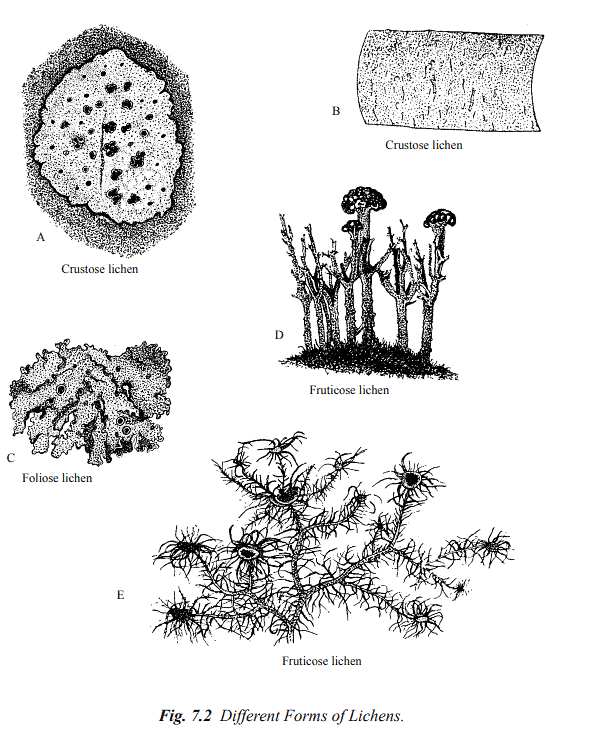
The simplest form of lichen thallus features loosely organized fungal hyphae and an algal component growing as powdery patches and is known as leprose lichen.
[The figure illustrates different forms of lichens: A—Crustose lichen (Haematomma sp.) growing on rock. B—Crustose lichen (Graphis scripta). C—Foliose lichen (Parmelia sp.) growing on bark; the dark cup-shaped structures are the apothecia (fruiting bodies). D—Fruticose lichen (Cladonia sp.) growing erect on the ground. E—Fruticose lichen (Usnea sp.) showing branched epiphytic thallus and disc-like apothecia.]
Internal Structure
Two primary types of lichen thalli can be identified based on their internal structure. In certain lichens, the fungi and photobionts (usually algae) are not restricted to a specific layer but are dispersed randomly throughout the thallus. This structural arrangement is termed "homiomerous."
The majority of lichens fall under the category of "heteromerous" or stratified thalli. In heteromerous lichens, the algal cells are organized into distinct layers.
These lichens exhibit internal differentiation into four main layers, as follows (see Figure 7.3):
(a) Upper Cortex: This layer consists of densely woven fungal hyphae with thick walls, often containing abundant gelatinous material. Interstitial spaces are typically absent in this layer.
(b) Algal Layer: In this layer, algal cells are surrounded by loosely packed fungal hyphae with thin cell walls.
(c) Medulla: Comprising the bulk of the thallus, the medulla consists of loosely packed fungal hyphae with thick cell walls.
(d) Lower Cortex: In some lichens, especially those with foliose forms (leaf-like), there may be a lower cortex composed of closely packed, dark-colored hyphae. This layer gives rise to rhizinae, specialized structures for attachment and nutrient absorption.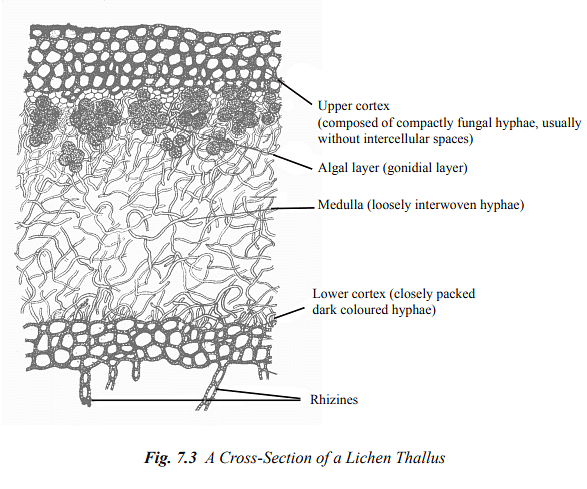
[Figure 7.3 illustrates the internal structure of lichen thalli.]
Special Structures of Lichen Thallus
- Cyphellae and Pseudocyphellae: Cyphellae and pseudocyphellae are tiny openings or fissures observed on the upper or lower layers of the lichen's cortex. These structures are crucial for facilitating the exchange of gases within the lichen thallus. Cyphellae possess a distinct rim and connect to the medulla of the lichen. They are only identified in the Sticta genus. On the other hand, pseudocyphellae are simpler in structure and lack a well-defined border.
[Figure 7.4 B provides an illustration of cyphellae in Sticta sp.]
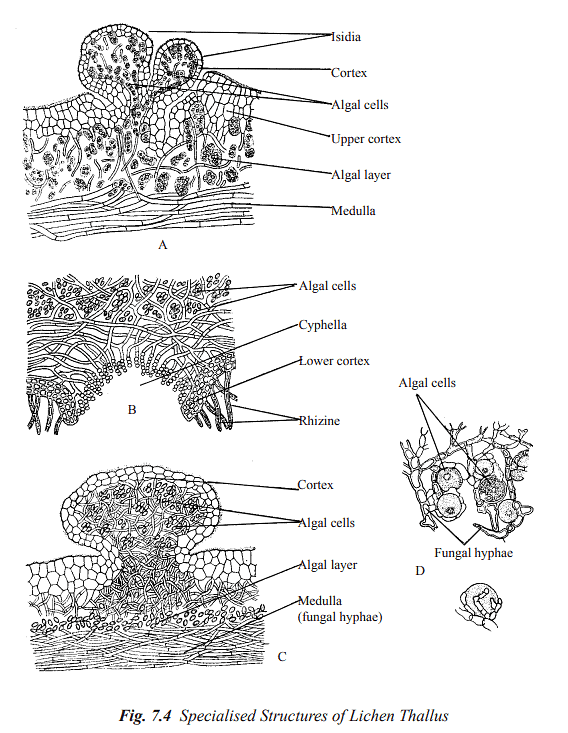
- Cephalodia: In numerous lichens, a fungus associates with two different photobionts, often a green alga and a cyanobacterium. In such instances, the cyanobacterial cells are confined to dark-colored, unique gall-like swellings called cephalodia, which are located on the upper surface of the lichen thallus. These cephalodia have the ability to fix nitrogen and continuously release it to the thallus. The fixed nitrogen is primarily utilized by the fungus. A notable example is Peltigera aphthosa.[Figure 7.4 C showcases cephalodia on the lichen's upper surface.]
- Soredia: Soredia are minuscule, rounded structures that form a powdery granular layer on the surface of the lichen thallus. They consist of a small cluster of algal cells closely surrounded by fungal hyphae. Wind disperses soredia, and they play a role in the vegetative reproduction of many lichen species. In certain lichens, soredia are produced within specialized pustule-like structures referred to as soralia (singular: soralium).
- Isidia: Isidia are diminutive, coral-like formations comprising both fungal hyphae and algal cells. They are typically found on the upper surface or edges of lichen lobes. When detached from the main lichen thallus, isidia have the capability to develop into new lichen thalli.
[Figure 7.4 A illustrates isidia on the lichen's surface.]
Reproduction
Lichens reproduce asexually and sexually.
Asexual Reproduction
It takes place by the following methods:
- Fragmentation: This is the commonest means of reproduction in lichens. Mere fragmentation of a thallus yields smaller pieces that can be transported to new habitats.
- Soredia: Many lichens, for example, Physcia spp., Ramalina spp., and Evernia produce specialsmallrounded granules or bud-like outgrowths, called soredia (sing.soredium) produced in pustule-like openings. Each soredium consists of one or more algal cells encircled by a few hyphal filaments. They are disseminated by wind or rain and on reaching a suitable substratum germinate to produce new thalli.
- Isidia: They are coral-like outgrowths on the uppersurface or margins of the lobes of lichens that have algal and fungal components walled inside a closed head-like structure. When detached, isidia can develop into new lichen thalli.
Sexual Reproduction
The method of sexual reproduction in lichens is contingent upon the specific type of fungus present in the lichen. Generally, the fungal component is an ascomycete, and as a result, ascocarps are produced. However, the reproductive capability of the photobiont is notably diminished in the lichenized state.
Types of Ascocarps: The ascocarp can assume the form of either an apothecium or a perithecium. In the case of an apothecium, its typical structure includes a layer of fertile tissues referred to as the hymenium or thecium. This hymenium comprises sterile, thread-like hyphae known as paraphyses, which are interspersed among sac-like structures called asci. These asci contain the ascospores.
[Figure 7.5 C provides an illustration of the apothecium's structure.]
Typically, each ascus (the sac-like structure) contains eight ascospores, although there may occasionally be one or two ascospores. Below the hymenium lies a layer of loosely packed fungal hyphae called the hypothecium or subhymenium.
Ascospore Germination and New Thallus Formation: The ascospores undergo germination to give rise to hyphal branches. When these branches make contact with a suitable algal cell, they initiate the development of a new lichen thallus.
Physiological Characteristics of the Lichen Thallus
Following are some physiological features of the composite lichen which suggest as to how they are adapted to the habitats in which they grow.
- Growth Rates: Lichens exhibit extremely slow growth rates. This sluggish growth allows them to thrive in environments where water and nutrient availability is limited.
- Adaptations to Fluctuating Moisture Conditions: Lichens are highly susceptible to desiccation in dry conditions but can rapidly absorb water when exposed to rain. They can also gradually absorb water vapor from humid atmospheres. For desert-dwelling lichen species, dew is a crucial source of water. Water availability is vital for their survival, and while fungi as a group can tolerate water stress to some extent, most algae are highly susceptible to drying out. The fungal medulla plays a crucial role by providing structural support for the autotrophic partner. This arrangement allows the autotrophic cells to be exposed in a thin layer to light. Additionally, the medulla contains a higher saturated water content compared to the rest of the thallus.
- Photosynthesis and Respiration: The rate of photosynthesis and respiration in lichens is closely tied to their water content. Due to the close association between the fungus and the autotrophic partner, some of the water becomes available to the autotroph, enabling it to photosynthesize even at lower water potentials.

- Absorption of Nutrients: Lichens are capable of absorbing substantial quantities of substances from their surroundings, with the primary responsibility for this uptake usually attributed to the fungal host.
- Lichen Substances or Lichen Acids: Lichens produce a diverse array of compounds known as lichen substances or lichen acids. These chemicals, primarily produced by the fungal partner, often exist as extracellular crystalline products. They are only synthesized when the fungus is in symbiosis. Lichen substances encompass various compounds, including fatty acids, lactones, polyketides, depsides, and anthraquinones. These metabolites play significant roles in processes like the weathering of rocks and the formation of soil (pedogenesis) through biogeochemical interactions. They are also known to function as chelating agents and antibiotics in certain soils. Many of these compounds appear to serve a defensive purpose, making lichens unappealing to grazing insects and mollusks, and inhibiting overgrowth by plants such as mosses. Pigmented compounds located in the upper cortex may contribute to protecting the symbiotic partners from intense light exposure.
Movement of Metabolites between the Partners in the Lichen Symbiosis
- Movement of Photosynthate from Symbiont to Fungus: The alga or cyanobacterium supplies the fungus with all its carbon requirements and with vitamins, such as biotin and thiamine. The carbohydrates, in whatever form they are released, become converted in the fungus into fungal carbohydrates, usually mannitol but sometimes into arabitol as well. The alga provides the fungus with necessary growth factors in the lichen.
- Movement of Fixed Nitrogen from Cyanobacterial Symbiont to Fungus: Many lichens with green algae possess cephalodia, small outgrowths on the surface, which usually contain a cyanobacterium. These fix nitrogen and constantly release it to the thallus where it is utilised almost exclusively by the fungus.
- Movement of substances from fungal partner to symbiont: It has long been claimed that the fungal host provides its symbionts with mineral nutrients. Lichen fungi have a marked ability to absorb and accumulate mineral ions from solution.
Nature of the Relationship between Algal and Fungal Partners in the Lichen Association
The precise nature of the relationship between the two partners in a lichen association remains a subject of extensive debate and is not yet fully comprehended.
- Mutualistic Symbiosis: The prevailing perspective posits that the association between the algal (or cyanobacterial) and fungal partners in lichens represents a form of mutualism, wherein all partners mutually benefit. In this context, the alga or cyanobacterium provides the fungus with essential carbohydrates and vitamins like biotin and thiamine. When the photosynthetic partner is a nitrogen-fixing cyanobacterium, it also supplies nitrogenous compounds to the fungus. Conversely, the fungus likely absorbs, stores, and furnishes water and necessary minerals to the alga or cyanobacterium. Additionally, it offers protection to the alga, shielding it from physical harm, and possibly from extreme temperatures. This particular relationship is often likened to a consortium, signifying a harmonious coexistence.
- Harustoria Formation: In certain lichens, the fungal partner produces specialized structures called haustoria, which penetrate the cells of the alga. While this might indicate a form of parasitism, it doesn't appear to severely harm the algal (or cyanobacterial) cells. They are neither killed nor significantly damaged. Consequently, some researchers characterize the relationship between fungi and algae (or cyanobacteria) as a balanced form of parasitism.
- Helotism Hypothesis: An alternate perspective suggests that the benefits of the lichen partnership predominantly favor the fungi. This viewpoint introduces the term "helotism" to describe such a relationship, where the fungus functions as the dominant organism, and the alga assumes a more subordinate role, akin to a "slave."
Resynthesis of the Lichen from the Partners
While it is relatively straightforward to isolate the majority of lichen algae (or cyanobacteria) and fungi into pure cultures, reconstituting lichens from these isolated partners has proven to be exceptionally challenging. In recent years, some associations have been created that resemble lichen structures, but they have not exhibited the distinctive characteristics of lichen bodies. Nevertheless, it has become evident that the most favorable conditions for lichen resynthesis involve an extremely nutrient-poor growth medium, a suitable substrate like rock, wood, or soil (rather than agar), and periodic, gradual cycles of drying and rehydration in the cultures.
Ecology of Lichens
- Lichens have the ability to thrive in extreme climatic conditions and impoverished habitats where most other organisms would struggle or perish. They can colonize exposed rocks in tropical, arctic, and alpine regions. Lichens are also capable of growing on dry, nutrient-poor tree bark and rocks in the supratidal zone, where they are consistently subjected to saltwater spray. Some lichen species are known to endure temperatures as low as -198ºC, while others inhabit rocks with temperatures exceeding 50ºC. Certain lichen species require alternating wet and dry periods for their normal growth. Although most lichens grow slowly, they have long lifespans.
- Lichens are often referred to as "pioneer" organisms because of their capacity to thrive in generally adverse conditions and their role in preparing an environment for other plants by facilitating rock weathering and the accumulation of organic matter. Lichens do not typically inhabit extremely hostile environments. Moreover, most lichens are exceptionally sensitive to air pollutants, and they are frequently employed as environmental pollution indicators due to this sensitivity.
Economic Importance of Lichens
- Pioneers of Vegetation: Lichens are recognized as "pioneer" organisms because they can prepare an environment for other plants by contributing to the weathering of rocks and the buildup of organic material. As rocks break down into soil particles, this creates a suitable substrate for the growth of mosses, followed by herbs, shrubs, and eventually trees.
- Lichens as a Food Source: Numerous small creatures like mites, slugs, snails, caterpillars, and termites feed on lichens such as Aspicilia calcarea and Lecanora saxicola. Some lichens, such as Cladonia rangiferina (reindeer moss), serve as important food sources for reindeer, caribou (musk ox), and other animals in the arctic and sub-arctic regions. Certain lichens contain a starch-like carbohydrate called lichenin, making them potentially useful for human consumption, although most are not very palatable. In Iceland, Cetraria islandica, known as Iceland moss, is used as a food source for sheep, cattle, and humans. To make it suitable for consumption, the lichen is soaked in a weak solution of sodium or potassium to neutralize lichen acids, dried, and then powdered. When boiled in water, the powder produces a jelly that is consumed with milk. Parmelia perlata has even been used as a curry ingredient by natives in the Bellary district of Madras. In Japan, a desert lichen called Endocarpon miniatum is used as a vegetable, and Lecanora is collected and eaten in Libya.
- Applications in Pharmacology and Medicine: Several lichen species were historically used in the treatment of various diseases, although they have largely been replaced by more effective drugs today. Lobaria pulmonaria and Cetraria islandica were employed to treat lung diseases, Xanthoria parientina for jaundice, and Peltigera canina (dog lichen) in cases of hydrophobia. Some lichen metabolites, like usnic acid, exhibit antibiotic properties against various fungi and bacteria, including human pathogens, and are used in commercially available antiseptic creams. Lichen substances are also known to possess anti-tumor activity, and some act as plant growth regulators.
- Lichens as a Source of Dyes: Lichens were historically used as a source of natural dyes, dating back to ancient Greek times. However, they have lost economic significance due to the widespread use of synthetic dyes. Erythrolitmin, a dye used in litmus paper to indicate pH changes, is extracted from various lichen species like Roccella, Parmelia, Evernia, and Ochrolechia.
- Use in Perfumery: Some lichen species contribute to the preparation of dhup hawan samagris and other perfumes. Evernia prunastri and Pseudovernia furfuracea (tree moss) are examples of lichens that provide an oily substance used in perfumes.
- Role in the Tanning Industry: Certain lichens, including Lobaria pulmonaria and Cetraria islandica, are utilized as tanning agents in the leather tanning industry.
- Harmful Effects of Lichen Substances: Specific lichen substances have been reported to cause contact dermatitis, a severe skin rash in humans.
|
179 videos|140 docs
|

|
Dec 22, 2024 Last updated |

|
Explore Courses for UPSC exam
|
|
Exam
,Structure and Reproduction Fungi & Lichens | Botany Optional for UPSC
,Structure and Reproduction Fungi & Lichens | Botany Optional for UPSC
,mock tests for examination
,Viva Questions
,practice quizzes
,study material
,Previous Year Questions with Solutions
,Summary
,ppt
,shortcuts and tricks
,Semester Notes
,past year papers
,MCQs
,Free
,Sample Paper
,Objective type Questions
,Important questions
,Structure and Reproduction Fungi & Lichens | Botany Optional for UPSC
,Extra Questions
,video lectures
;






Structure and Reproduction Fungi & Lichens Free PDF Download
Importance of Structure and Reproduction Fungi & Lichens
Structure and Reproduction Fungi & Lichens Notes
Structure and Reproduction Fungi & Lichens UPSC Questions
Study Structure and Reproduction Fungi & Lichens on the App
|
© EduRev
|
Education Revolution
|



|




