Fertilization | Zoology Optional Notes for UPSC PDF Download

Introduction
Fertilization is a intricate process involving the merging of male and female gametes, accompanied by the fusion of their cytoplasm. The commencement of fertilization is marked by the sperm's proximity to the egg and concludes with the fusion of the egg and sperm pronuclei.
Mechanism of fertilization
The process of fertilization completes in the following five stages:
- Encounter of spermatozoa and ova
- Approach of the sperm to the egg.
- Acrosome reaction and penetration.
- Activation of ovum.
- Migration of pronuclei and Amphimixis
Encounter of Spermatozoa and Ova
One of the challenges in sexual reproduction is facilitating the encounter between spermatozoa and ova in a fluid medium, allowing individual sperm to reach the surface of eggs at the right moment. The key requirements for this encounter are a fluid medium for fertilization and the delivery of a significant quantity of sperm in proximity to the number of mature eggs at the appropriate time.
There are two reported types of fertilization based on the location and nature of the fluid medium: external fertilization and internal fertilization.
- External Fertilization:
- Occurs in a liquid medium outside the bodies of parent animals.
- Among freshwater animals like fishes, amphibians, and freshwater invertebrates, the timing of egg laying by the female and sperm shedding by the male is highly specific. The short lifespan of sperm necessitates direct delivery to individual eggs immediately after laying.
- Marine forms release eggs and sperm freely into the surrounding water. The extended time interval between egg laying and sperm shedding can be weeks or months due to the saltwater serving as a vital physiological medium for gametes.
- Sperm movement in a liquid medium during external fertilization is random, with collisions between sperm and eggs occurring by chance. The high number of sperm produced by males and the relatively larger size of eggs contribute to successful encounters.
- Internal Fertilization:
- Takes place inside the female's body.
- In oviparous forms like reptiles and birds, eggs are enclosed in impermeable egg membranes. In ovoviviparous and viviparous animals, eggs are retained within the maternal body during development.
- Spermatozoa are internally delivered to the female's body through copulatory mechanisms or male intromittant organs. Fertilization may occur in different locations, such as the lower or upper parts of the oviduct, ovarian follicles in viviparous fishes, and eutherian mammals.
- In internal fertilization, the movement of spermatozoa from the deposition site to the fertilization site often relies on the active swimming of spermatozoa themselves, passive transport through muscular contractions of the female tract, and counter-currents in the cilia-propelled, backward-flowing liquid content of the tract.
Approach of the sperm to the egg
The sperm's approach to the egg involves various methods:
- Chemotaxis:
- Chemotaxis refers to the sperm's ability to detect differences in the concentration of substances released by the egg into the water, moving from lower to higher concentrations. This mechanism is observed in plants, mosses, ferns, coelenterates, and fishes.
- In certain instances, such as in the hydroid Campanularia, chemical substances from the female gonangia attract sperm towards the opening of the theca.
- Fertilizin and Anti-fertilizin Interactions:
- Eggs and sperm of some animals, including mammals, exhibit chemical attraction. Substances released by eggs cause agglutination or clumping of sperm on the egg surface.
- Fertilizin and anti-fertilizin, found in eggs and sperm, respectively, play a direct role in the interaction between the egg and sperm. Fertilizin is mainly located in the egg's plasma membrane, while in sea urchin eggs, it is produced by the jelly layer surrounding the egg.
- Fertilizin is a gel consisting of glycoprotein or mucopolysaccharide, with species-specific variations. It has a large molecular weight and can attach to multiple spermatozoa, binding them together. Sperm plasma membrane contains antifertilizin, a species-specific protein.
- Capacitation:
- In mammals, ejaculated or epididymal sperm do not undergo acrosome reaction unless they spend some time in the female genital duct.
- Capacitation involves the gradual removal of coating substances on the sperm surface, particularly those on the acrosome, exposing receptor sites. This process occurs over varying time spans in different species.
- Capacitation is crucial as it allows sperm to recognize signals from the ovulated egg. It is induced by factors in the female genital tract and is not strictly species-specific.
- The process is significant for preventing premature release of hydrolytic and proteolytic enzymes in the acrosome, ensuring the safety of the male genital tract. Capacitation enables the acrosomes to release enzymes, facilitating sperm penetration of the egg during fertilization.
Acrosome Reaction and Penetration
- Most animal eggs are surrounded by protective layers, such as egg membranes, gelatinous layers, or follicle cells, outside the plasma membrane. These layers act as barriers to sperm penetration, preventing fertilization by more than one sperm or by sperm from different species. When sperm attaches to the egg surface, it becomes immobile, and its penetration through egg membranes and plasma membranes is facilitated by the physico-chemical activity of the sperm acrosome.
- The sperm acrosome produces enzymatic proteins known as sperm lysins, which vary across animal groups. In some cases, such as with echinoderm eggs, the dissolution of the egg envelopes may be triggered by the acidification of seawater due to carbon dioxide produced by the sperm during respiration. In eggs with thick envelopes, like those of fishes and insects, sperm must penetrate through a specific canal called the micropyle.
- In mammals, eggs released from the ovary are encased in a layer of follicular cells called the corona radiata. This layer is held together by an adhesive substance called hyaluronic acid. The corona radiata acts as a barrier that sperm must penetrate to reach the egg's plasma membrane. The sperm acrosome produces an enzyme, hyaluronidase, which dissolves the adhesive substance, dispersing the cells of the corona radiata.
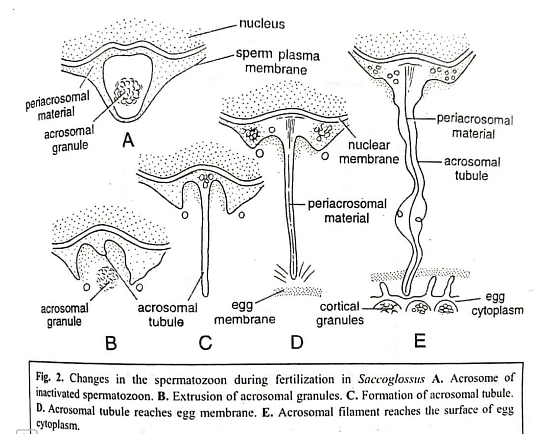
- The penetration of membranous barriers is mediated not only by lytic agents from the sperm acrosome but also by morphological changes in the acrosome, forming an acrosomal filament that aids penetration into the egg interior. This process has been well-documented in echinoderms, annelids, and Saccoglossus (hemichordata).
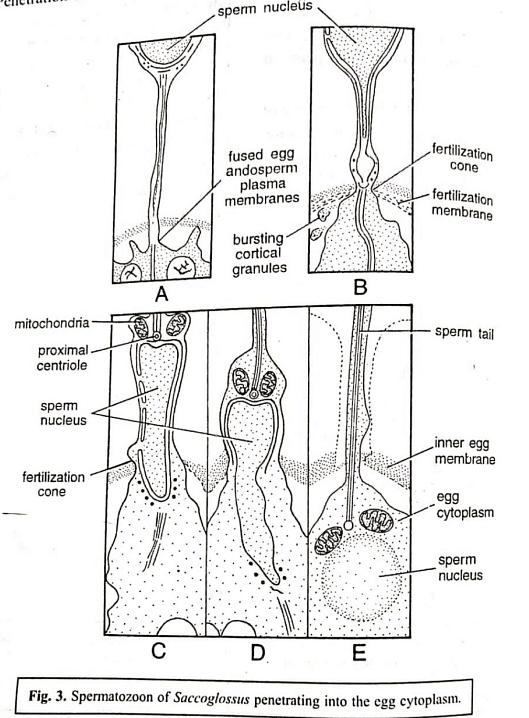
- The acrosome reaction and penetration in Saccoglossus involve several events, including the bursting of the acrosome, release of lytic enzymes, formation of an acrosomal tubule, eversion of the acrosomal membrane, fusion of the acrosomal tubule with the egg membrane, and passage of sperm contents. Similar events occur in other animals during sperm penetration, with the activation of sperm involving the rupturing of the acrosome and the formation of acrosomal filaments or tubules. Mammalian sperm, although possessing an acrosome, does not develop acrosomal filaments; instead, the acrosomal and egg membranes dissolve at the point of contact, allowing the sperm to enter the egg's interior.
Activation of Ovum
Activation of the ovum refers to the process in fertilization where an egg transitions from an inactive state to an active state, initiating development. Upon contact between the acrosomal tubule of the sperm and the egg's plasma membrane, fusion occurs, creating a continuous mosaic membrane, resulting in the formation of a single cell known as the zygote. This moment triggers essential changes in the egg's cytoplasm, including the formation of the fertilization cone, cortical reactions leading to the fertilization membrane, and metabolic activation.
A. Fertilization Cone Formation:
Right after the acrosomal filament of the sperm makes contact with the egg's surface, the egg's cytoplasm protrudes at the point of contact, forming a structure known as the fertilization cone. The fertilization cone can take various shapes, including a simple conical protrusion, several irregular pseudopodium-like processes, or even a cytoplasmic cylinder extending along the acrosomal filament or tubule. Regardless of its specific form, the fertilization cone envelops the sperm and subsequently retracts.
B. Cortical Reactions and Formation of Fertilization Membrane
Even before the fertilization cone is formed and the sperm penetrates into the interior of egg, a chain of physico-chemical reactions is set in the cortex. All these reactions are collectively called cortical reaction. These reactions may differ from one group of animals to another, but in most groups, the cortical reactions lead to the formation of fertilization membrane outside the egg plasma membrane. This membrane blocks the entry of the late arriving spermatozoa in the egg interior and thus avoids polyspermy.
The process of cortical and fertilization membrane formation in different geoups of animals is as under:
Sea Urchins: In sea urchins, as soon as the apical end of acrosomal tubule touches the surface of egg from the site of contact, a wave like colour change from yellow to white travels rapidly arounf=d the egg cortex and is immediately followed by the elevation of fertilization cone from the egg surface and the formation of fertilization membrane around the egg plasma membrane. Electron micrographs of Sea Urchins’ unfertilized eggs show that the egg cortex is bound by two membranes: an outer vitelline membrane and an inner plasma membrane.
Beneath the plasma membrane occurs a layer of cortical granules. A fertilization membrane is formed in the following stages:
The outer vitelline membrane separated from the plasma membrane undergoes expansion and becomes the outer layer of the fertilization membrane. The cortical granules explode and release the following three components:
- Dark, denser, lamellar and folded bodies (i.e., electron opaque material) of the cortical granules unfold and fuse with the inner side of the already elevated membrane, the vitelline membrane.
- Globules (hemisphere bodies), fuse together and build up a new surface of the viscous hyaline layer, just at the outer side of the egg plasma membrane. The hyaline layer adhered closely to the surface of the egg and during cleavage, it helps to keep the blastomeres together.
- The liquefied component of the cortical granules fills the perivitelline space between the new egg surface and now the completed and elevated fertilization membrane. It contains mucopolysaccharides and abundant water.
All these structures, namely vitelline membrane and contents of cortical granules, thus form the fertilization membrane, which is much thicker (up to 900Å) and stronger. In vertebrates, the changes which occur in the cortex are similar to sea urchins with some minor exceptions. e.g., the unfertilized eggs of some mammals (man, rabbit, etc.) have cortical granules. In them, the sperm penetration is not followed by the formation of fertilization membrane but, the cortical granules burst open and release their contents into the perivitelline space, i.e. the space between the egg plasma membrane zona pellucida. In urodel amphibians and some mammals, which lack cortical granules, neither any cortical reaction nor fertilization membrane formation occurs.
C. Metabolic Activation: After the sperm penetrates the unfertilized egg, a series of cytoplasmic reaction is initiated.
Following metabolic changes occur in the egg during fertilization:
- Changes in plasma membrane: The permeability of plasma membrane increases for the molecules of water and certain other chemicals like ethylene glycol, phosphate, K + , etc. During fertilization, the electrical potential of plasma membrane becomes more positive and gradually becomes more negative. The change in the electrical potential of the membrane is governed by the unequal distribution of chloride ions (Cl - ). Besides this, a plasma membrane enzyme adenyl cyclase becomes activated at the time of fertilization and it starts the formation of a chemical molecule 3' 5' cyclic AMP, which is supposed to activate most of the metabolic reactions in a fertilized egg.
- Ionic changes: Certain intracellular changes occur in the concentration of cations, especially those of sodium, potassium and calcium. The change in calcium ion concentration in a fertilized egg has great significance in the metabolic activation of the egg.
- Changes in the rate of respiration: In a fertilized egg, the rate of respiration either increases (e.g., sea urchins) or decreases (e.g., Chaetopterus) or remains static (e.g., Bufo). There appears to be a relation between the post-fertilization oxygen consumption and the stage of maturation of the egg at fertilization. Because at the time of fertilization the sea urchin egg has completed maturation, the egg of Bufo is at the second maturation division stage and the egg of Chaetopterus is at the first maturation division stage. The increased oxygen consumption is related with the oxidation of glycogen and other food stuff of the egg and synthesis of numerous ATP molecules.
- Co-enzyme changes: In a fertilized egg, inter-conversion of coenzyme, NAD into another co-enzyme NADP and also NADPH due to phosphorylation of the NAD in the presence of an enzyme NAD Kinase takes place.

There is ample evidence that NAD kinase enzyme although present in the unfertilized egg, is in inactive state. It is activated only at the time of fertilization. The increased NADP and NADPH contents may initiate many synthetic pathways of fertilized eggs. - Changes in the rate of protein synthesis: The cytoplasm of a mature unfertilized egg contains complete machinery for protein synthesis, such as DNA molecules, tRNA, MRNA, ribosomes and proteolytic enzymes required during protein synthesis, but none or very little protein synthesis occurs because the mRNA of unfertilized egg remains 'masked'. There is evidence that during later phases of oogenesis some inhibitor or repressor proteins are manufactured in sea urchin’s egg which inactivate chromosomal genes, mRNA molecules, ribosomes etc. During fertilization, there is an increase in proteolytic activity of the egg immediately following the penetration of sperm, which removes these inhibitor proteins from them and unmasks the mRNA and active protein synthesis is started. In the egg of frog, however, the rate of protein synthesis is increased quite early at the stage of ovulation itself.
- Initiation of mitosis: The initiation of mitosis for cleavage is the most significant aspect of egg activation. The initiation of mitosis occurs because (i) the rate of DNA synthesis increases with great pace immediately after fertilization, (ii) the unfertilized egg cytoplasm although possesses a centriole, yet this centriole is incapable of division and also to form a mitotic spindle. Thus, sperm stimulates the first mitotic division (cleavage) of fertilized egg by contributing its centriole to the egg. In some animals after fertilization, a considerable amount of acid is formed during the first several months.
Theories of Activation
The repressor theory of activation proposes that during the egg's maturation, energy-yielding systems are blocked, inhibiting metabolic and genetic activities. Accumulation of inhibitory substances is suggested, and the key event in fertilization is the removal of these repressors, unleashing cytoplasmic metabolic activities and genetic systems.
Repressor theory of activation
In a recent study by Monroy (1965), compelling arguments were presented suggesting that the activation of the egg by the sperm occurs at the molecular level. According to Monroy, the energy-yielding systems within the egg are obstructed during maturation, inhibiting various reactions, including metabolic activities in the egg cytoplasm, which demand a substantial amount of energy. The likely explanation is the accumulation of inhibitory substances during egg maturation, and Monroy identified one such inhibitor.
Monroy's repressor theory has gained support from contemporary embryologists like Tomkins et al. (1969), Metafora et al. (1971), Berrill (1971), and D. Epel (1973). Substantial evidence now exists, indicating that repressor substances produced by the egg in the later stages of maturation inhibit both the metabolic activities in the cytoplasm and the genetic activities of the nucleus. The crucial step in fertilization, therefore, involves removing these repressors, simultaneously unleashing cytoplasmic metabolic activities and activating the nuclear genetic system.
Recent investigations have provided insights into the initial events that activate the egg. Following the acrosome reaction, there is a surge in Ca++ levels and an elevation in pH within the sperm. This localized increase in cytoplasmic Ca++ at the sperm-egg contact site appears sufficient to initiate the exocytosis of cortical granules and the formation of the fertilization membrane. Subsequently, a propagating wave of Ca++ is released from intracellular stores, accompanied by an increase in cytoplasmic pH. These two components, Ca++ and pH, seem to be the primary regulators triggering various cytoskeletal changes during fertilization.
Components of Sperm in the Egg Interior
Variations are evident across different animal groups regarding the extent to which sperm components enter the egg during fertilization. Typically, the sperm nucleus, peri-acrosomal material, proximal centriole, and mitochondria enter the egg. The sperm's plasma membrane becomes integrated into the egg's plasma membrane. In mammals, the entire sperm structure, including the head, middle piece, and tail, penetrates the egg cytoplasm. In echinoderms, the sperm tail remains outside the vitelline membrane, while in Nereis, only the sperm nucleus and proximal centriole enter the egg cytoplasm. There is no conclusive evidence that any sperm component, except the nucleus and centrosome, actively contributes to subsequent development. Although mitochondria have been observed in the egg cytoplasm, their duration of existence remains unknown.
In vertebrates, eggs typically complete their first meiotic division in the ovary, reaching the metaphase stage of the second meiotic division. Further progression is halted at this stage, and ovulation occurs, potentially leading to fertilization. The extrusion of the second polar body only happens if the egg is fertilized by a sperm.
In ascidians, eggs only reach the metaphase of the first meiotic division when they become ripe. If fertilized, the egg then completes the first reduction division and undergoes the second meiotic division.
Migration of Pronuclei and Amphimixis
Upon sperm penetration into the egg cytoplasm, the sperm nucleus retains a compact form, with its mitochondria and centriole situated behind it. To initiate amphimixis, the sperm nucleus undergoes two crucial activities: (i) transformation into a pronucleus and (ii) migration to the amphimixis site. As the sperm nucleus moves inward from the fertilization cone site, it undergoes a 180°C rotation, positioning its mitochondria and centriole at the forefront. Simultaneously, the sperm nucleus swells, and its densely packed chromatin becomes finely granular, eventually adopting a vesicular appearance resembling the interphase nucleus, referred to as the male pronucleus.
Concurrently, within the egg cytoplasm, a sperm aster forms around the proximal centriole of the sperm. As the male pronucleus develops and moves toward the amphimixis site, the sperm aster appears to guide it. The amphimixis site is typically near the center for microlecithal and mesolecithal eggs or at the animal pole's active cytoplasm center for microlecithal and telolecithal eggs. The movement of the sperm pronucleus may be accompanied by cortical and subcortical cytoplasm, with pigmented granules marking its trajectory, known as the penetration path, especially in heavily pigmented amphibian eggs. Some researchers suggest a directed movement due to a chemotaxic effect of chemicals released by the female pronucleus. Deviations from the penetration path result in a new route known as the copulation path in some cases, while in others, the penetration and copulation paths remain identical.
Before amphimixis, the egg nucleus undergoes changes similar to the sperm nucleus. After the second meiotic division, the haploid egg nucleus is found near the egg surface as vesicles called karyomeres. These karyomeres fuse to form a female pronucleus, which swells, increases in volume, and becomes vesicular, migrating toward the amphimixis site.
Amphimixis, the fusion of male and female pronuclei, involves the breakdown of nuclear membranes at the contact point. Their contents unite within a common nuclear membrane. As the first cleavage of the fertilized egg approaches, the nuclear membrane dissolves, and chromosomes from both maternal and paternal origins align on the equator of the achromatic spindle.
|
198 videos|351 docs
|
FAQs on Fertilization - Zoology Optional Notes for UPSC
| 1. What is the process of fertilization? |  |
| 2. How does fertilization occur? | |
| 3. What are the theories of activation in fertilization? | |
| 4. How does fertilization lead to the development of a new individual? | |
| 5. What are the factors that can affect fertilization? | |



Exam
,Important questions
,Previous Year Questions with Solutions
,shortcuts and tricks
,Fertilization | Zoology Optional Notes for UPSC
,video lectures
,ppt
,Semester Notes
,Objective type Questions
,Extra Questions
,practice quizzes
,Viva Questions
,Fertilization | Zoology Optional Notes for UPSC
,MCQs
,past year papers
,Sample Paper
,Summary
,mock tests for examination
,Fertilization | Zoology Optional Notes for UPSC
,study material
,Free
;






Fertilization Free PDF Download
Importance of Fertilization
Fertilization Notes
Fertilization UPSC Questions
Study Fertilization on the App
|
© EduRev
|
Education Revolution
|



|





